

This is a preprint.
Diffusing protein binders to intrinsically disordered proteins
- PMID: 39071267
- PMCID: PMC11275890
- DOI: 10.1101/2024.07.16.603789
Diffusing protein binders to intrinsically disordered proteins
Update in
-
Diffusing protein binders to intrinsically disordered proteins.Nature. 2025 Aug;644(8077):809-817. doi: 10.1038/s41586-025-09248-9. Epub 2025 Jul 30. Nature. 2025. PMID: 40739343 Free PMC article.
Abstract
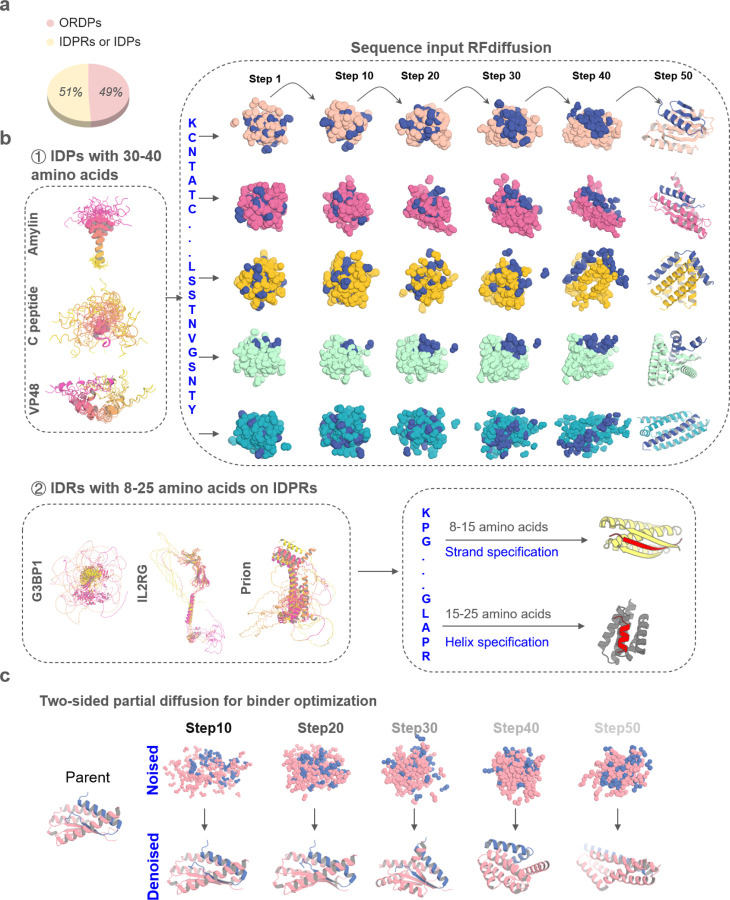
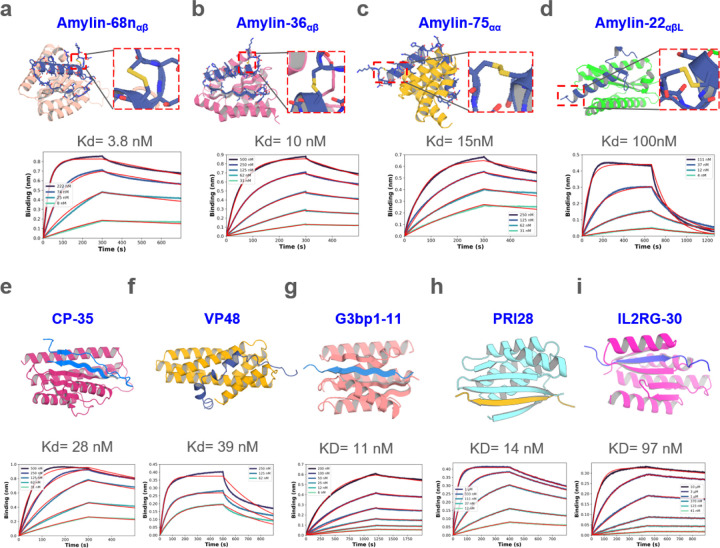
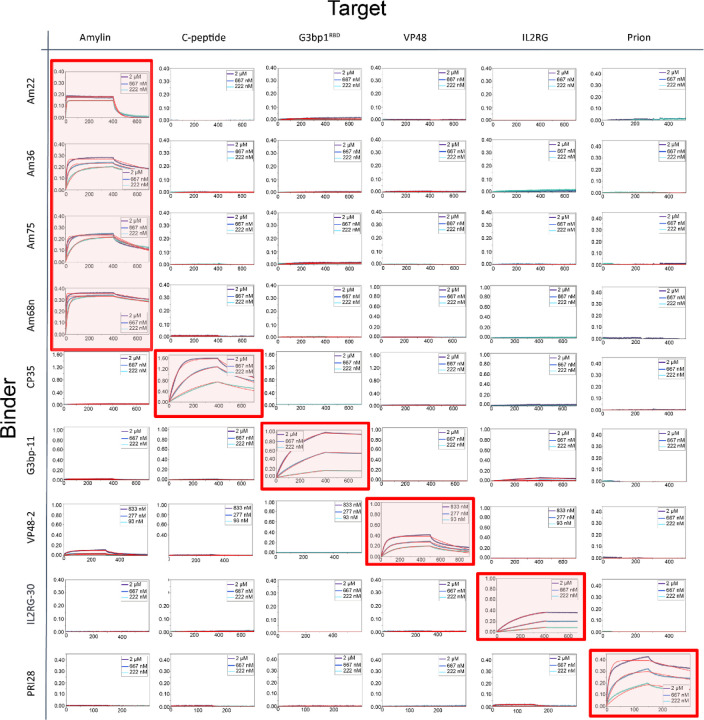
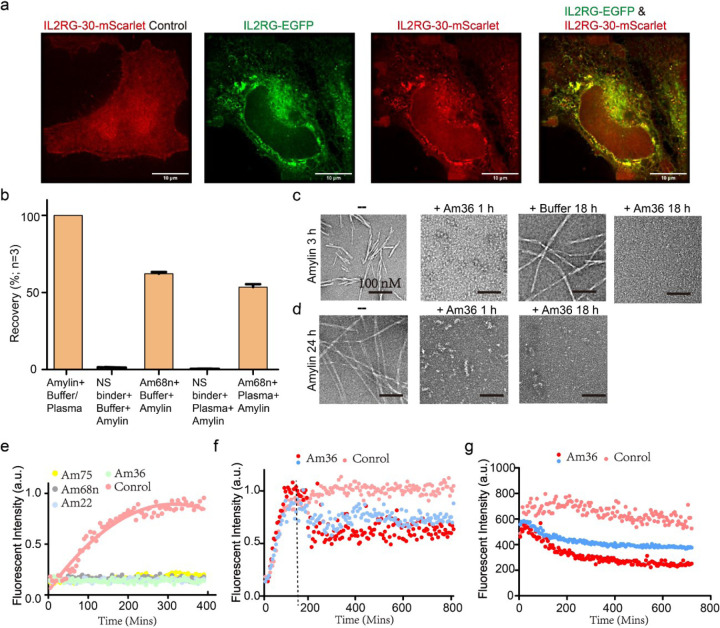
Proteins which bind intrinsically disordered proteins (IDPs) and intrinsically disordered regions (IDRs) with high affinity and specificity could have considerable utility for therapeutic and diagnostic applications. However, a general methodology for targeting IDPs/IDRs has yet to be developed. Here, we show that starting only from the target sequence of the input, and freely sampling both target and binding protein conformation, RFdiffusion can generate binders to IDPs and IDRs in a wide range of conformations. We use this approach to generate binders to the IDPs Amylin, C-peptide and VP48 in a range of conformations with Kds in the 3 -100nM range. The Amylin binder inhibits amyloid fibril formation and dissociates existing fibers, and enables enrichment of amylin for mass spectrometry-based detection. For the IDRs G3bp1, common gamma chain (IL2RG) and prion, we diffused binders to beta strand conformations of the targets, obtaining 10 to 100 nM affinity. The IL2RG binder colocalizes with the receptor in cells, enabling new approaches to modulating IL2 signaling. Our approach should be widely useful for creating binders to flexible IDPs/IDRs spanning a wide range of intrinsic conformational preferences.
Keywords: Amyloid fibril dissociation; Intrinsically disordered protein; RFdiffusion; Rosetta; deep learning; diagnostics; intrinsically disordered region; protein design.
Figures

References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous