

This is a preprint.
Cytoplasmic ribosomes on mitochondria alter the local membrane environment for protein import
- PMID: 39071314
- PMCID: PMC11275913
- DOI: 10.1101/2024.07.17.604013
Cytoplasmic ribosomes on mitochondria alter the local membrane environment for protein import
Update in
-
Cytoplasmic ribosomes on mitochondria alter the local membrane environment for protein import.J Cell Biol. 2025 Apr 7;224(4):e202407110. doi: 10.1083/jcb.202407110. Epub 2025 Mar 6. J Cell Biol. 2025. PMID: 40047641 Free PMC article.
Abstract
Most of the mitochondria proteome is nuclear-encoded, synthesized by cytoplasmic ribosomes, and targeted to mitochondria post-translationally. However, a subset of mitochondrial-targeted proteins is imported co-translationally, although the molecular mechanisms governing this process remain unclear. We employ cellular cryo-electron tomography to visualize interactions between cytoplasmic ribosomes and mitochondria in Saccharomyces cerevisiae. We use surface morphometrics tools to identify a subset of ribosomes optimally oriented on mitochondrial membranes for protein import. This allows us to establish the first subtomogram average structure of a cytoplasmic ribosome on the surface of the mitochondria in the native cellular context, which showed three distinct connections with the outer mitochondrial membrane surrounding the peptide exit tunnel. Further, this analysis demonstrated that cytoplasmic ribosomes primed for mitochondrial protein import cluster on the outer mitochondrial membrane at sites of local constrictions of the outer and inner mitochondrial membrane. Overall, our study reveals the architecture and the spatial organization of cytoplasmic ribosomes at the mitochondrial surface, providing a native cellular context to define the mechanisms that mediate efficient mitochondrial co-translational protein import.
Figures

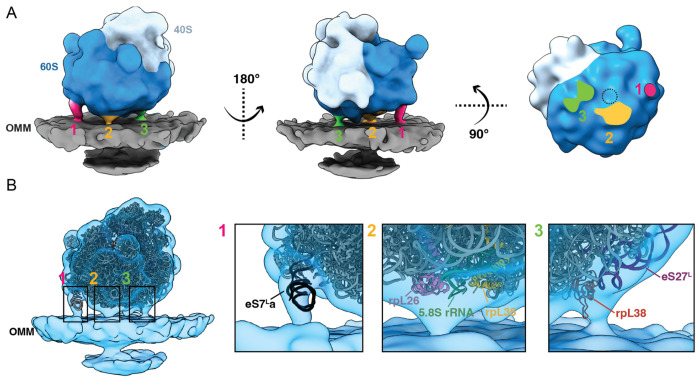
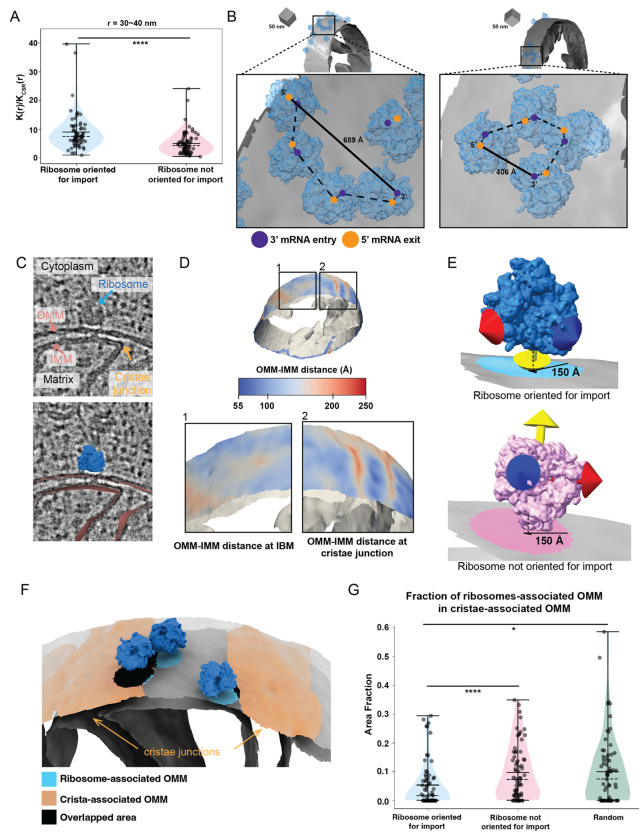

References
-
- Avendaño-Monsalve M. C., Ponce-Rojas J. C. and Funes S. (2020). “From cytosol to mitochondria: the beginning of a protein journey.” Biol Chem 401(6-7): 645–661. - PubMed
-
- Aviram N. and Schuldiner M. (2017). “Targeting and translocation of proteins to the endoplasmic reticulum at a glance.” J Cell Sci 130(24): 4079–4085. - PubMed
-
- Becker T., Bhushan S., Jarasch A., Armache J.-P., Funes S., Jossinet F., Gumbart J., Mielke T., Berninghausen O., Schulten K., Westhof E., Gilmore R., Mandon E. C. and Beckmann R. (2009). “Structure of monomeric yeast and mammalian Sec61 complexes interacting with the translating ribosome.” Science (New York, N.Y.) 326(5958): 1369–1373. - PMC - PubMed
-
- Brandt F., Carlson L. A., Hartl F. U., Baumeister W. and Grünewald K. (2010). “The three-dimensional organization of polyribosomes in intact human cells.” Mol Cell 39(4): 560–569. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources