

This is a preprint.
Variants in the DDX6-CXCR5 autoimmune disease risk locus influence the regulatory network in immune cells and salivary gland
- PMID: 39071447
- PMCID: PMC11275775
- DOI: 10.1101/2023.10.05.561076
Variants in the DDX6-CXCR5 autoimmune disease risk locus influence the regulatory network in immune cells and salivary gland
Update in
-
Variants in the DDX6-CXCR5 autoimmune disease risk locus influence the regulatory network in immune cells and salivary gland.Ann Rheum Dis. 2025 Sep;84(9):1512-1527. doi: 10.1016/j.ard.2025.04.023. Epub 2025 May 30. Ann Rheum Dis. 2025. PMID: 40447495
Abstract
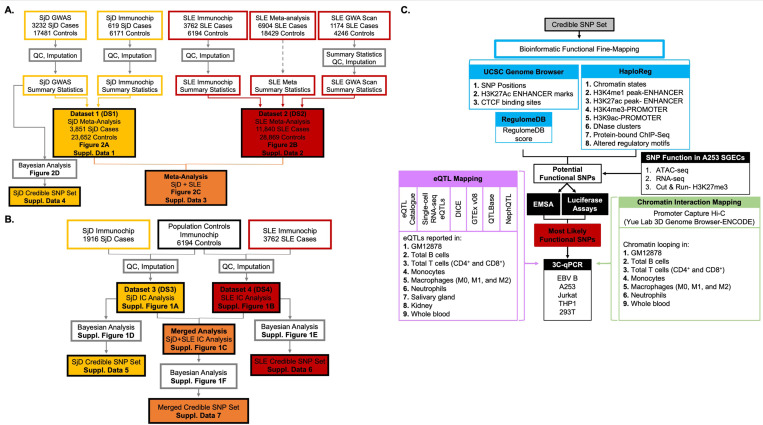
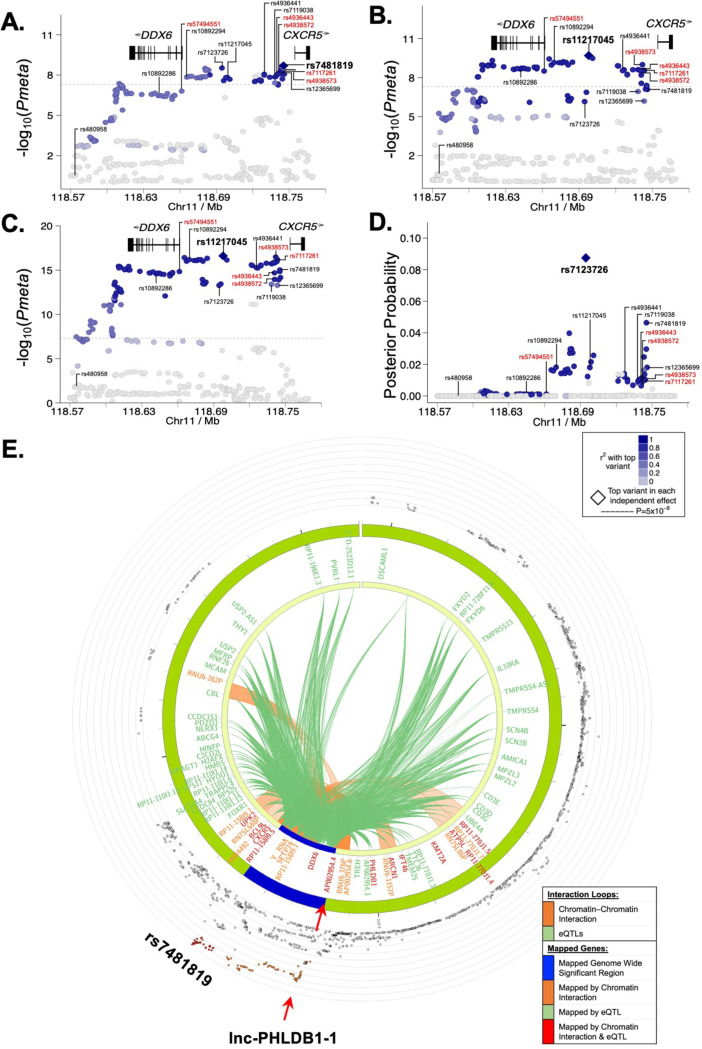
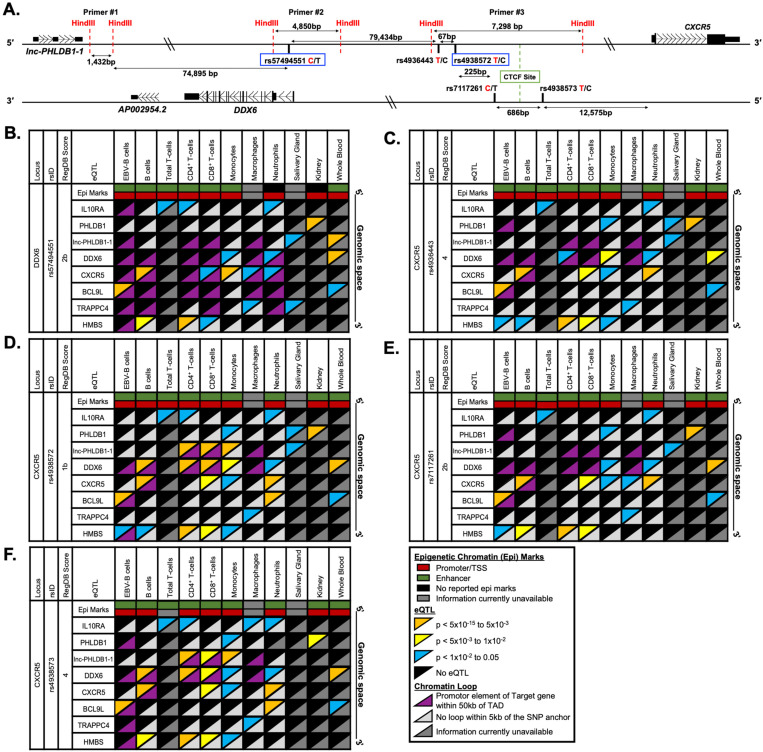
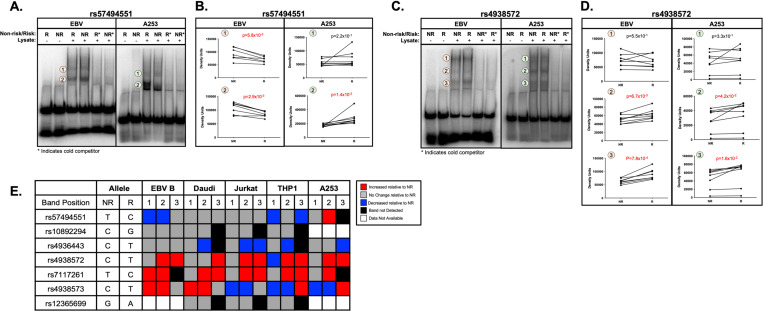
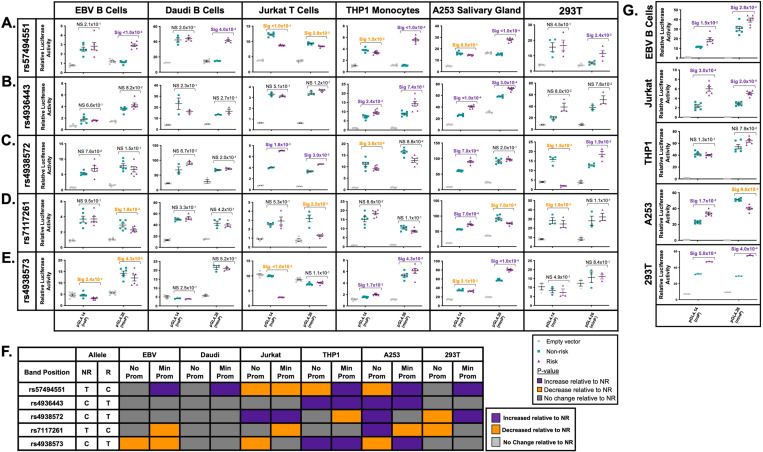
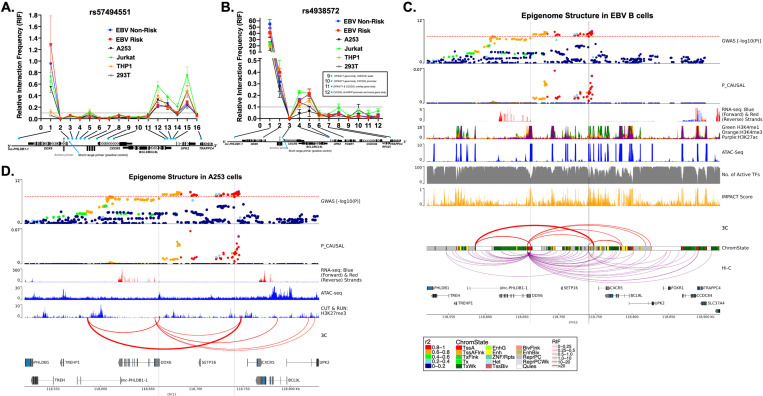
Fine mapping and bioinformatic analysis of the DDX6-CXCR5 genetic risk association in Sjögren's Disease (SjD) and Systemic Lupus Erythematosus (SLE) identified five common SNPs with functional evidence in immune cell types: rs4938573, rs57494551, rs4938572, rs4936443, rs7117261. Functional interrogation of nuclear protein binding affinity, enhancer/promoter regulatory activity, and chromatin-chromatin interactions in immune, salivary gland epithelial, and kidney epithelial cells revealed cell type-specific allelic effects for all five SNPs that expanded regulation beyond effects on DDX6 and CXCR5 expression. Mapping the local chromatin regulatory network revealed several additional genes of interest, including lnc-PHLDB1-1. Collectively, functional characterization implicated the risk alleles of these SNPs as modulators of promoter and/or enhancer activities that regulate cell type-specific expression of DDX6, CXCR5, and lnc-PHLDB1-1, among others. Further, these findings emphasize the importance of exploring the functional significance of SNPs in the context of complex chromatin architecture in disease-relevant cell types and tissues.
Keywords: C-X-C motif chemokine receptor 5; CXCR5; DDX6; DEAD-Box helicase 6; SLE; Sjögren’s Disease; chromatin regulatory network; lnc-PHLDB1-1; systemic lupus erythematosus.
Conflict of interest statement
Competing Interests C.J.L.* and A.D.F. have an active collaborative research agreement with Janssen. E.B. has an active research collaboration with Pfizer. T.M. is employed as medical solutions lead in rheumatology at UCB. R.H.S. is a consultant for Jansen Pharmaceuticals. S.J.B. provided consultancy services for Abbvie, BMS, Galapagos, Iqvia, J&J, Kiniksa, and Novartis in 2020–2021. L.R. provided consultancy services for AstraZeneca. B.M.W. has active collaborative research agreements with Astellas Bio and Pfizer, Inc. M.R. received grants from Amgen, AstraZeneca, Bristol Myers-Squibb, Novartis, and Servier for clinical trials in Sjögren’s Syndrome and SLE. All other authors have reported that they have no competing interests to report.
Figures
References
Publication types
Grants and funding
- RC4 AG039029/AG/NIA NIH HHS/United States
- U01 HG004446/HG/NHGRI NIH HHS/United States
- Z01 DE000704/ImNIH/Intramural NIH HHS/United States
- R01 AR065953/AR/NIAMS NIH HHS/United States
- HHSN268201200008C/HL/NHLBI NIH HHS/United States
- HHSN268200782096C/HG/NHGRI NIH HHS/United States
- R01 AR071947/AR/NIAMS NIH HHS/United States
- R01 AR073855/AR/NIAMS NIH HHS/United States
- R33 AR076803/AR/NIAMS NIH HHS/United States
- N01 DE032636/DE/NIDCR NIH HHS/United States
- RC2 AG036495/AG/NIA NIH HHS/United States
- P50 CA093459/CA/NCI NIH HHS/United States
- HHSN268201200008I/HL/NHLBI NIH HHS/United States
- R21 AR079089/AR/NIAMS NIH HHS/United States
- R01 CA133996/CA/NCI NIH HHS/United States
- P50 CA097007/CA/NCI NIH HHS/United States
- R61 AR076803/AR/NIAMS NIH HHS/United States
- P50 AR060804/AR/NIAMS NIH HHS/United States
- UM1 AI144292/AI/NIAID NIH HHS/United States
- R01 AR071410/AR/NIAMS NIH HHS/United States
- U01 DE028891/DE/NIDCR NIH HHS/United States
- HHSN268201100011I/HL/NHLBI NIH HHS/United States
- HHSN268201100011C/HL/NHLBI NIH HHS/United States
- R01 ES011740/ES/NIEHS NIH HHS/United States
- P30 AR070155/AR/NIAMS NIH HHS/United States
- P30 AR073750/AR/NIAMS NIH HHS/United States
- R03 DE029800/DE/NIDCR NIH HHS/United States
- R01 AR074310/AR/NIAMS NIH HHS/United States
LinkOut - more resources
Full Text Sources