

Template-assisted covalent modification underlies activity of covalent molecular glues
- PMID: 39075252
- PMCID: PMC11582070
- DOI: 10.1038/s41589-024-01668-4
Template-assisted covalent modification underlies activity of covalent molecular glues
Abstract
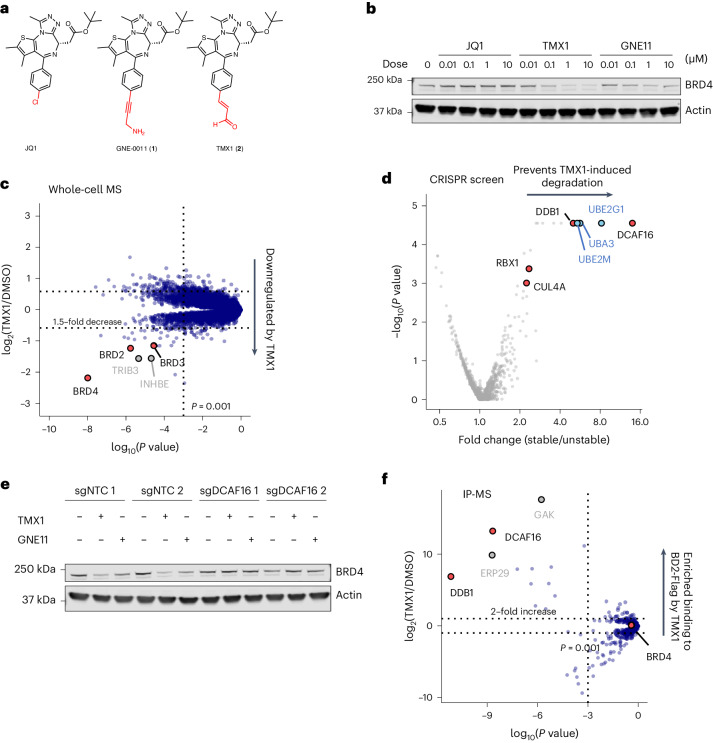
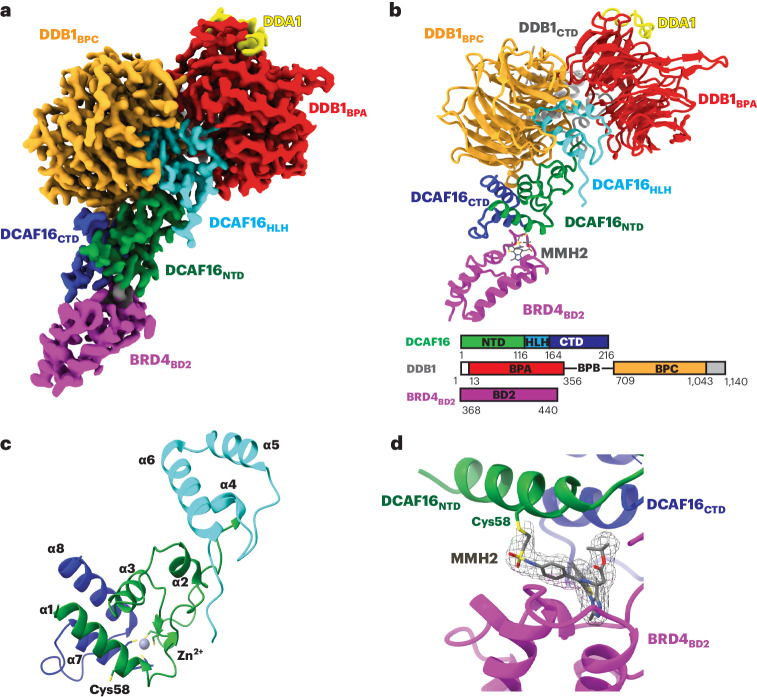
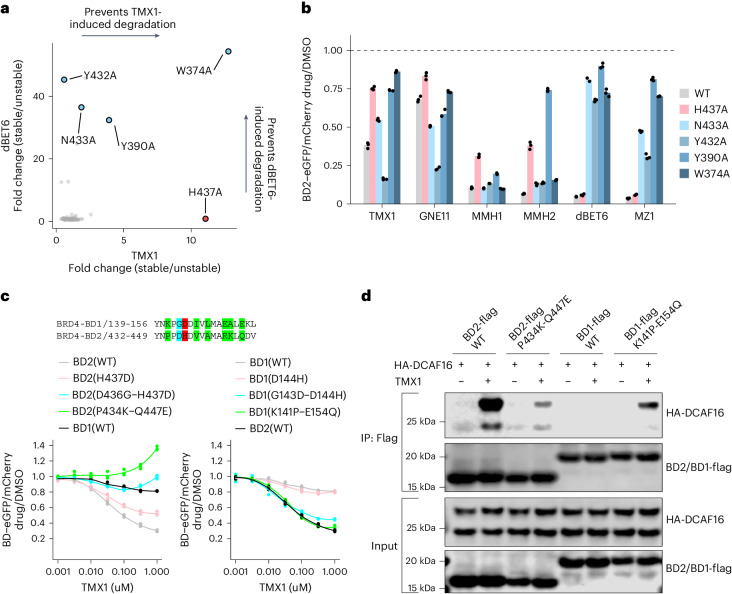
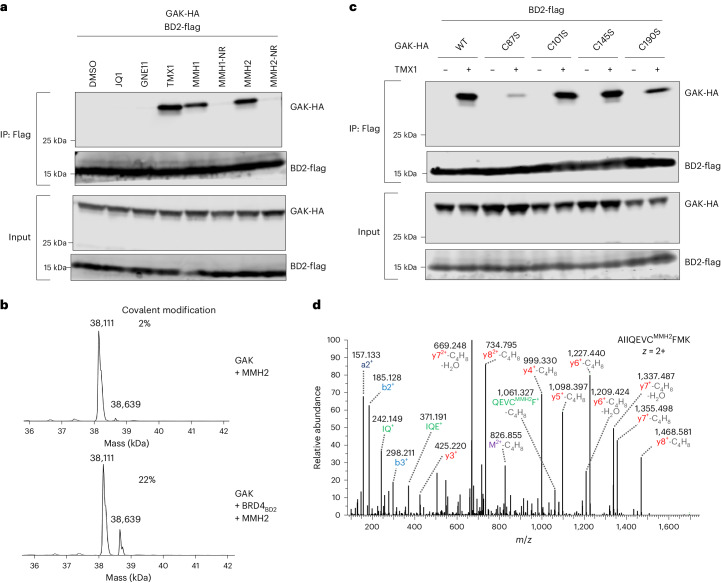
Molecular glues are proximity-inducing small molecules that have emerged as an attractive therapeutic approach. However, developing molecular glues remains challenging, requiring innovative mechanistic strategies to stabilize neoprotein interfaces and expedite discovery. Here we unveil a trans-labeling covalent molecular glue mechanism, termed 'template-assisted covalent modification'. We identified a new series of BRD4 molecular glue degraders that recruit CUL4DCAF16 ligase to the second bromodomain of BRD4 (BRD4BD2). Through comprehensive biochemical, structural and mutagenesis analyses, we elucidated how pre-existing structural complementarity between DCAF16 and BRD4BD2 serves as a template to optimally orient the degrader for covalent modification of DCAF16Cys58. This process stabilizes the formation of BRD4-degrader-DCAF16 ternary complex and facilitates BRD4 degradation. Supporting generalizability, we found that a subset of degraders also induces GAK-BRD4BD2 interaction through trans-labeling of GAK. Together, our work establishes 'template-assisted covalent modification' as a mechanism for covalent molecular glues, which opens a new path to proximity-driven pharmacology.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: B.L.E. has received research funding from Celgene, Deerfield, Novartis and Calico Life Sciences and consulting fees from AbbVie. He is a member of the scientific advisory board (SAB) for and a shareholder of Neomorph, Inc., TenSixteen Bio, Skyhawk Therapeutics and Exo Therapeutics. E.S.F. is a founder, SAB member and equity holder of Civetta Therapeutics, Lighthorse Therapeutics, Proximity Therapeutics and Neomorph, Inc. (board of directors). He is an equity holder in and SAB member for Avilar Therapeutics and Photys Therapeutics and a consultant to Novartis, Sanofi, EcoR1 Capital and Deerfield. The Fischer laboratory receives or has received research funding from Deerfield, Novartis, Ajax, Interline and Astellas. N.S.G. is a founder, SAB member and equity holder in Syros, C4, Allorion, Lighthorse, Voronoi, Inception, Matchpoint, CobroVentures, GlaxoSmithKline, Larkspur (board member), Shenandoah (board member) and Soltego (board member). The Gray laboratory receives or has received research funding from Novartis, Takeda, Astellas, Taiho, Jansen, Kinogen, Arbella, Deerfield, Springworks, Interline and Sanofi. M.S. has received research funding from Calico Life Sciences. K.A.D. receives or has received consulting fees from Kronos Bio and Neomorph, Inc. J.Q. is an equity holder of Epiphanes and Talus Bioscience and receives or has received research funding from Novartis. J.A.M. is a founder and equity holder of and advisor to Entact Bio, serves on the SAB of 908 Devices and receives or has received sponsored research funding from Vertex, AstraZeneca, Taiho, Springworks, TUO Therapeutics and Bruker. Y.-D.L. is currently employed by Leerink Partners. M.W.M. is currently employed by Novartis Venture Fund. K.P. is currently employed by AbbVie. R.J.L. is currently employed by Flagship Pioneering. B.J.G. is currently employed by Blueprint Medicines. I.Y. is currently employed by Matchpoint Therapeutics. The remaining authors declare no competing interests.
Figures












Update of
-
Template-assisted covalent modification of DCAF16 underlies activity of BRD4 molecular glue degraders.bioRxiv [Preprint]. 2023 Feb 15:2023.02.14.528208. doi: 10.1101/2023.02.14.528208. bioRxiv. 2023. Update in: Nat Chem Biol. 2024 Dec;20(12):1640-1649. doi: 10.1038/s41589-024-01668-4. PMID: 36824856 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
- R01CA262188/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R35 CA253125/CA/NCI NIH HHS/United States
- S10 OD028697/OD/NIH HHS/United States
- P50 CA206963/CA/NCI NIH HHS/United States
- R01 CA219850/CA/NCI NIH HHS/United States
- R21CA247671/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R01CA233800/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- T32 GM144273/GM/NIGMS NIH HHS/United States
- R01CA219850/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- U24 DK116204/DK/NIDDK NIH HHS/United States
- K99CA287069/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- K00CA253754/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- U24DK116204/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- 1S10OD028697-01/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R01 HL082945/HL/NHLBI NIH HHS/United States
- K00 CA253754/CA/NCI NIH HHS/United States
- P01 CA066996/CA/NCI NIH HHS/United States
- R21 CA247671/CA/NCI NIH HHS/United States
- RR220012/Cancer Prevention and Research Institute of Texas (Cancer Prevention Research Institute of Texas)
- K99 CA287069/CA/NCI NIH HHS/United States
- R01 CA233800/CA/NCI NIH HHS/United States
- P50CA206963/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R01 CA262188/CA/NCI NIH HHS/United States
- R35CA253125/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R01HL082945/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- P01CA066996/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- T32 CA236754/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
