

TMEM9 activates Rab9-dependent alternative autophagy through interaction with Beclin1
- PMID: 39078420
- PMCID: PMC11335249
- DOI: 10.1007/s00018-024-05366-1
TMEM9 activates Rab9-dependent alternative autophagy through interaction with Beclin1
Abstract
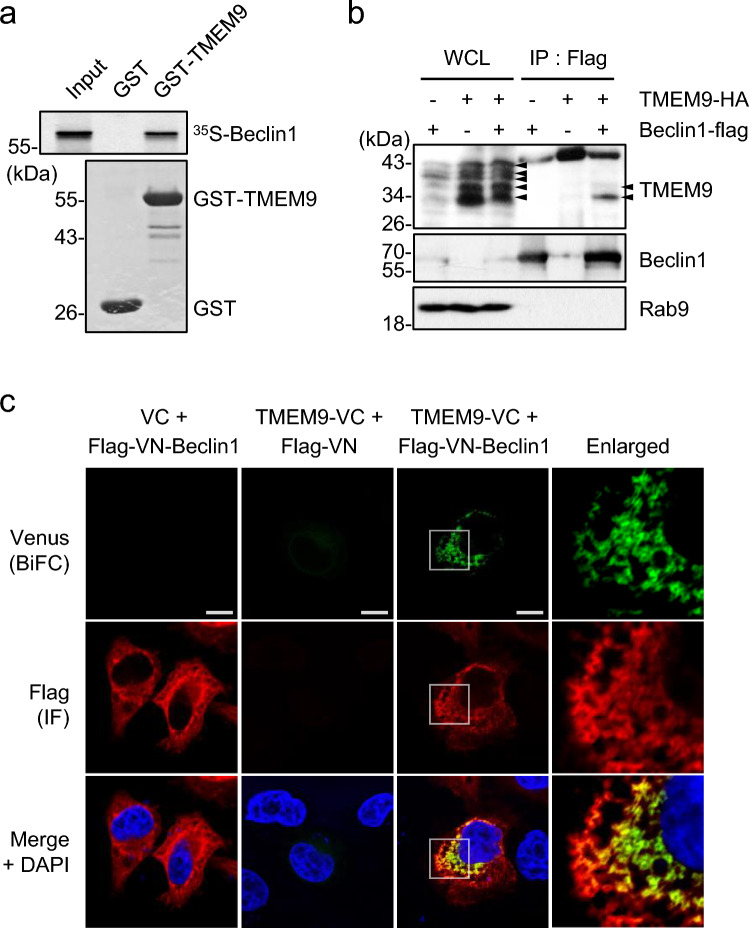
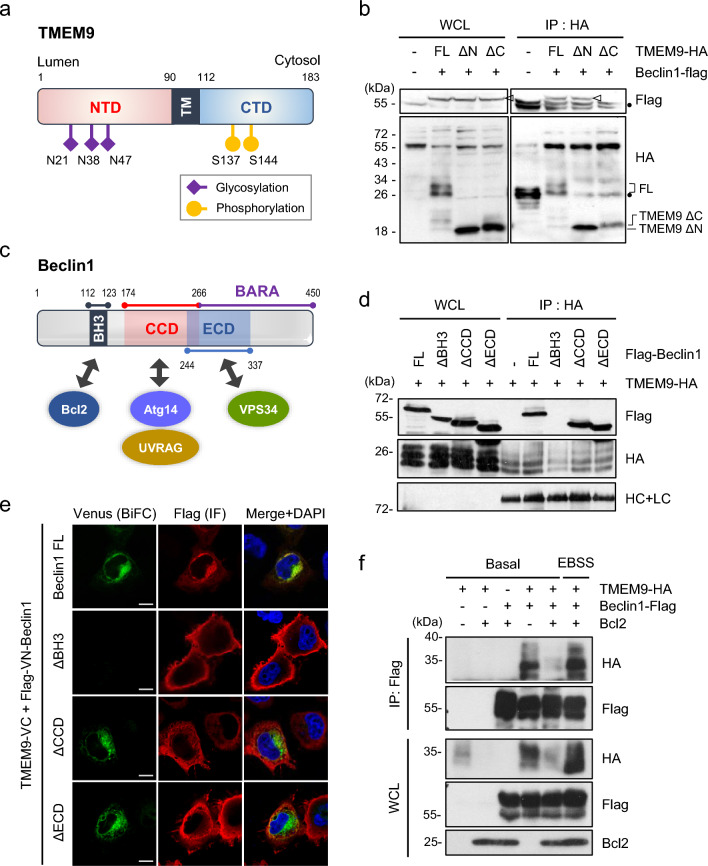
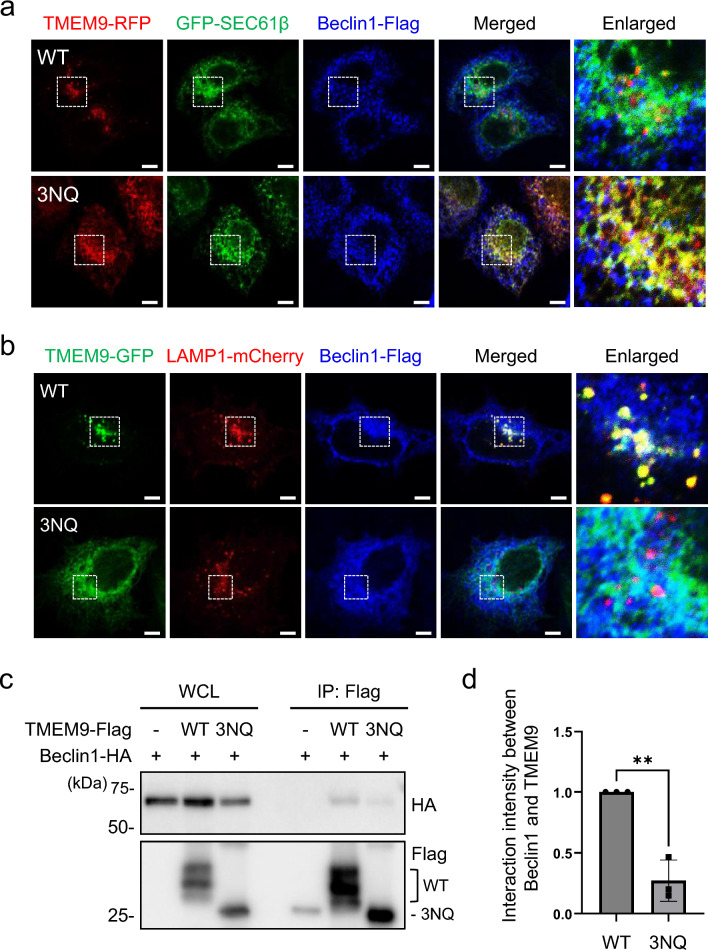
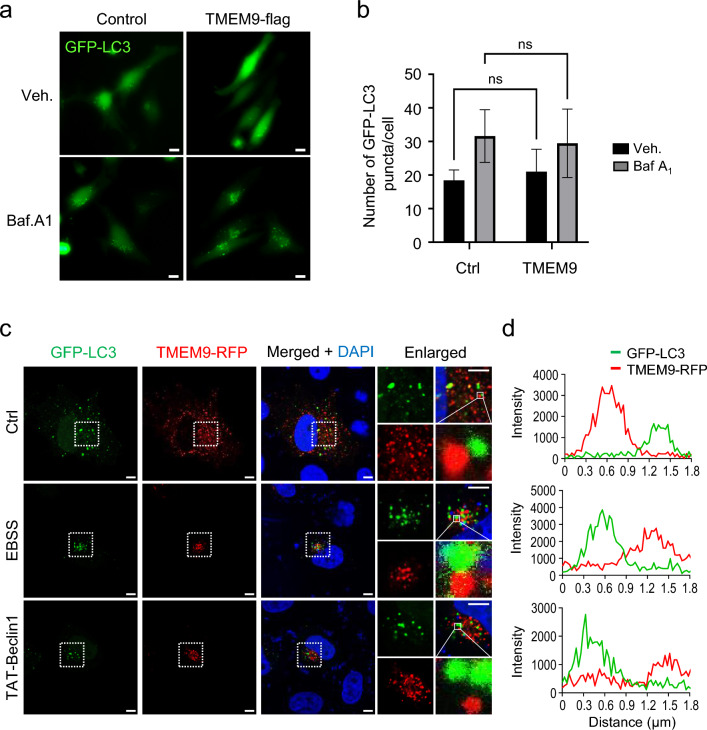
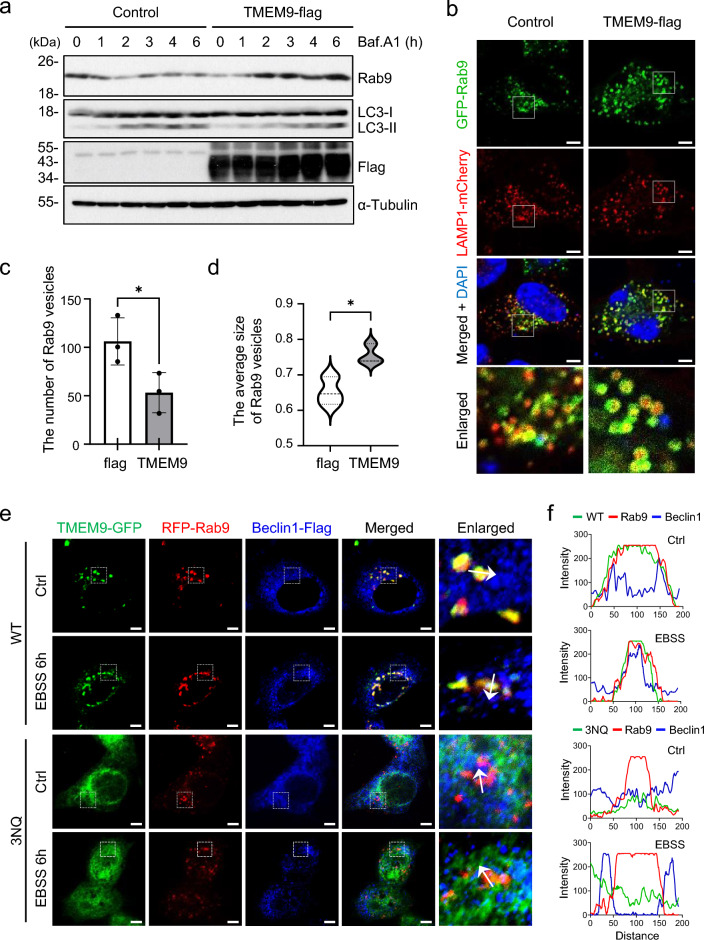
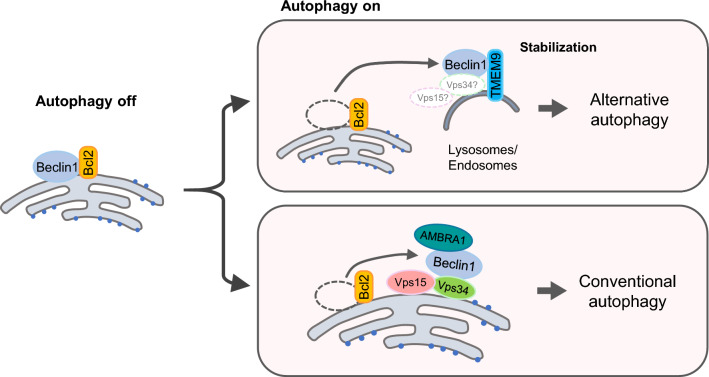
Transmembrane protein 9 (TMEM9) is a transmembrane protein that regulates lysosomal acidification by interacting with the v-type ATPase complex. However, the role of TMEM9 in the lysosome-dependent autophagy machinery has yet to be identified. In this study, we demonstrate that the lysosomal protein TMEM9, which is involved in vesicle acidification, regulates Rab9-dependent alternative autophagy through its interaction with Beclin1. The cytosolic domain of TMEM9 interacts with Beclin1 via its Bcl-2-binding domain. This interaction between TMEM9 and Beclin1 dissociates Bcl-2, an autophagy-inhibiting partner, from Beclin1, thereby activating LC3-independent and Rab9-dependent alternative autophagy. Late endosomal and lysosomal TMEM9 apparently colocalizes with Rab9 but not with LC3. Furthermore, we show that multiple glycosylation of TMEM9, essential for lysosomal localization, is essential for its interaction with Beclin1 and the activation of Rab9-dependent alternative autophagy. These findings reveal that TMEM9 recruits and activates the Beclin1 complex at the site of Rab9-dependent autophagosome to induce alternative autophagy.
Keywords: Alternative autophagy; Beclin1; Rab9; TMEM9.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no conflicting interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
