

Missense variants in CMS22 patients reveal that PREPL has both enzymatic and nonenzymatic functions
- PMID: 39078710
- PMCID: PMC11385081
- DOI: 10.1172/jci.insight.179276
Missense variants in CMS22 patients reveal that PREPL has both enzymatic and nonenzymatic functions
Abstract
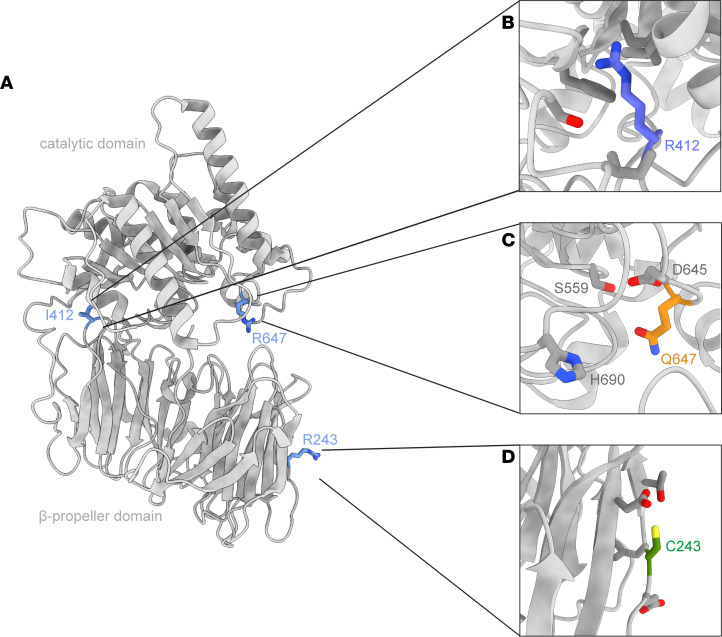
Congenital myasthenic syndrome-22 (CMS22, OMIM 616224) is a rare genetic disorder caused by deleterious genetic variation in the prolyl endopeptidase-like (PREPL) gene. Previous reports have described patients with deletions and nonsense variants in PREPL, but nothing is known about the effect of missense variants in the pathology of CMS22. In this study, we have functionally characterized missense variants in PREPL from 3 patients with CMS22, all with hallmark phenotypes. Biochemical evaluation revealed that these missense variants do not impair hydrolase activity, thereby challenging the conventional diagnostic criteria and disease mechanism. Structural analysis showed that the variants affect regions most likely involved in intraprotein or protein-protein interactions. Indeed, binding to a selected group of known interactors was differentially reduced for the 3 variants. The importance of nonhydrolytic functions of PREPL was investigated in catalytically inactive PREPL p.Ser559Ala cell lines, which showed that hydrolytic activity of PREPL is needed for normal mitochondrial function but not for regulating AP1-mediated transport in the transgolgi network. In conclusion, these studies showed that CMS22 can be caused not only by deletion and truncation of PREPL but also by missense variants that do not necessarily result in a loss of hydrolytic activity of PREPL.
Keywords: Endocrinology; Genetic variation; Genetics.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
