

Deletion of RFX6 impairs iPSC-derived islet organoid development and survival, with no impact on PDX1+/NKX6.1+ progenitors
- PMID: 39080045
- PMCID: PMC11604831
- DOI: 10.1007/s00125-024-06232-2
Deletion of RFX6 impairs iPSC-derived islet organoid development and survival, with no impact on PDX1+/NKX6.1+ progenitors
Abstract
Aims/hypothesis: Homozygous mutations in RFX6 lead to neonatal diabetes accompanied by a hypoplastic pancreas, whereas heterozygous mutations cause MODY. Recent studies have also shown RFX6 variants to be linked with type 2 diabetes. Despite RFX6's known function in islet development, its specific role in diabetes pathogenesis remains unclear. Here, we aimed to understand the mechanisms underlying the impairment of pancreatic islet development and subsequent hypoplasia due to loss-of-function mutations in RFX6.
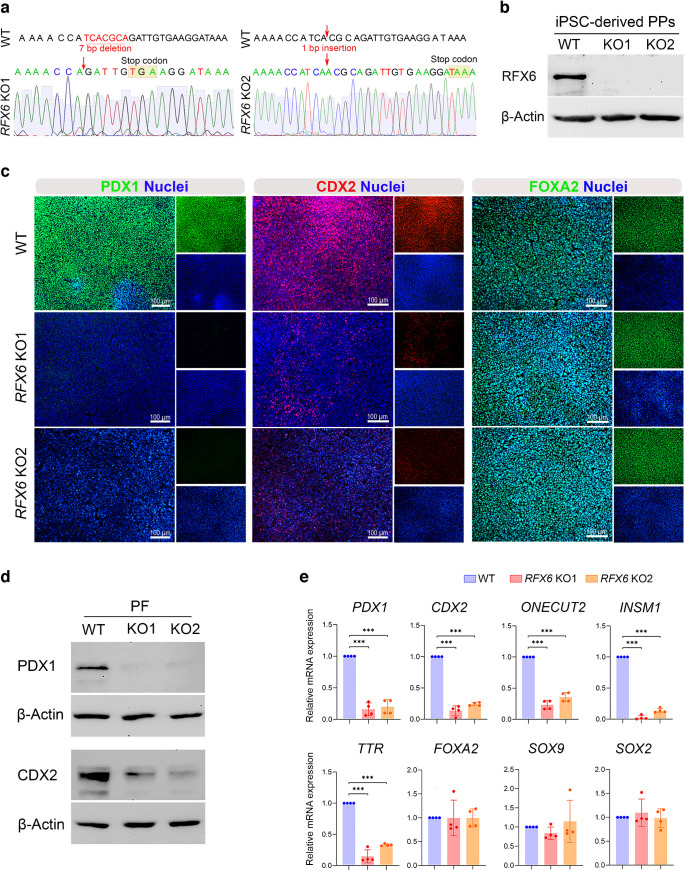
Methods: We examined regulatory factor X6 (RFX6) expression during human embryonic stem cell (hESC) differentiation into pancreatic islets and re-analysed a single-cell RNA-seq dataset to identify RFX6-specific cell populations during islet development. Furthermore, induced pluripotent stem cell (iPSC) lines lacking RFX6 were generated using CRISPR/Cas9. Various approaches were then employed to explore the consequences of RFX6 loss across different developmental stages. Subsequently, we evaluated transcriptional changes resulting from RFX6 loss through RNA-seq of pancreatic progenitors (PPs) and endocrine progenitors (EPs).
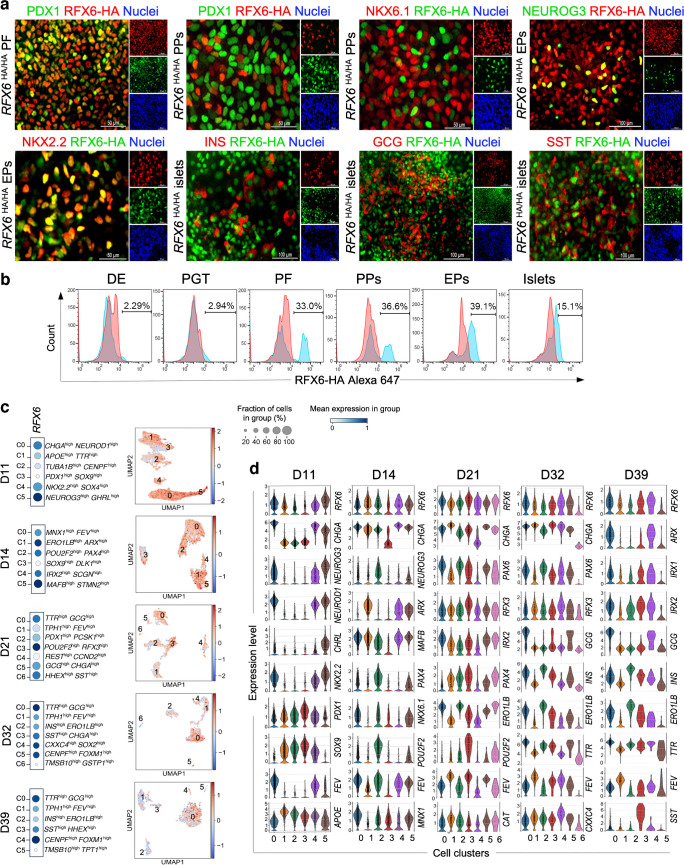
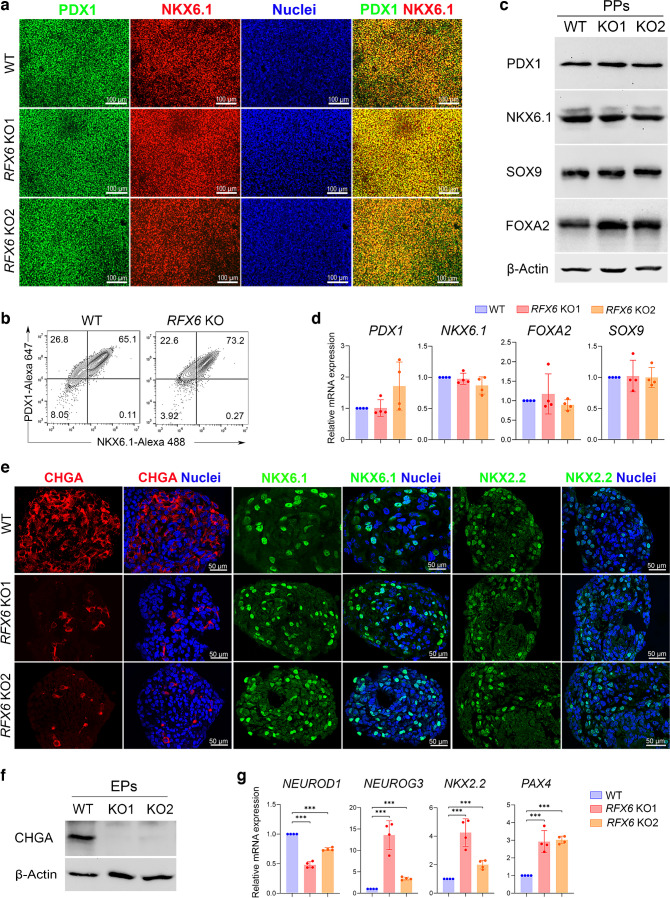
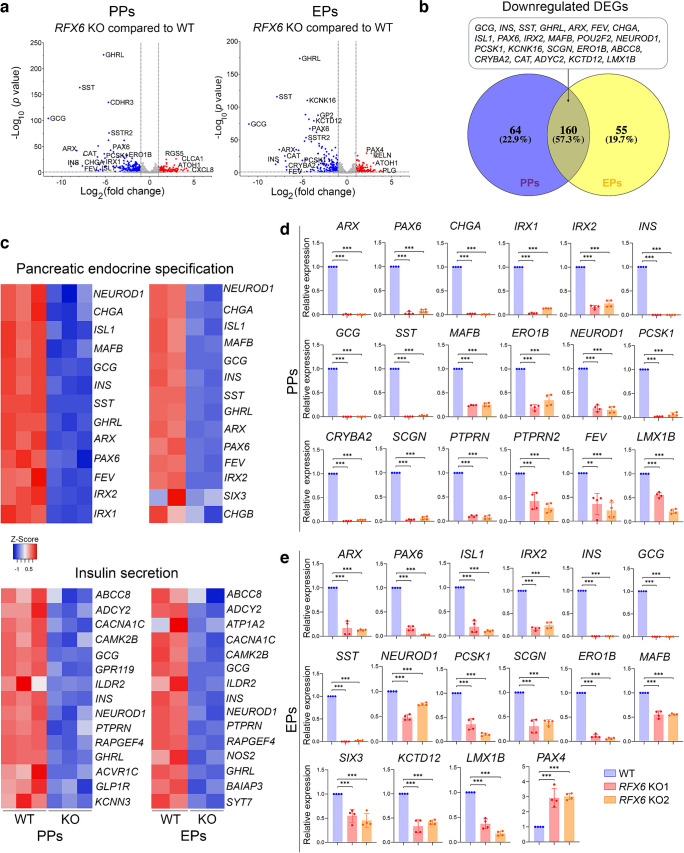
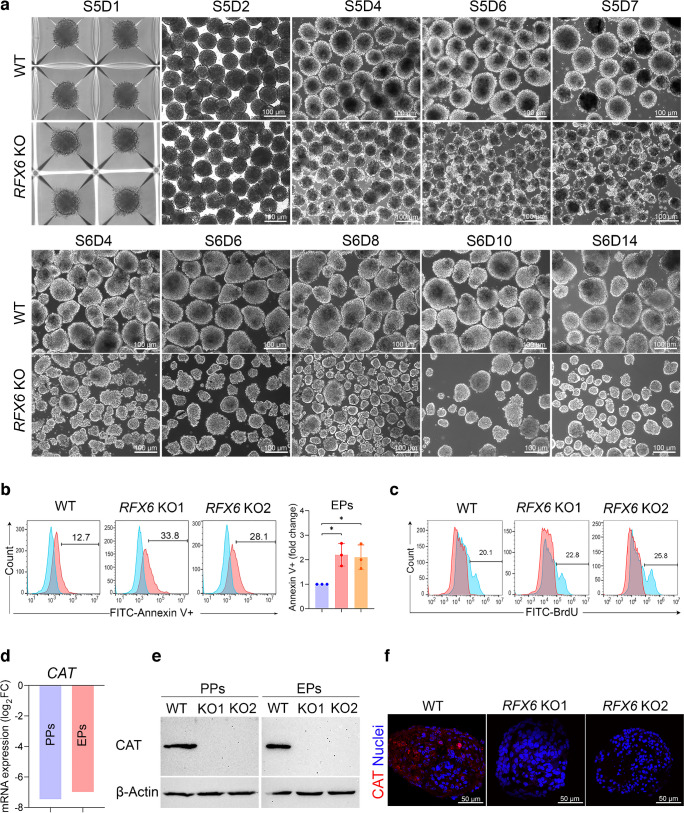
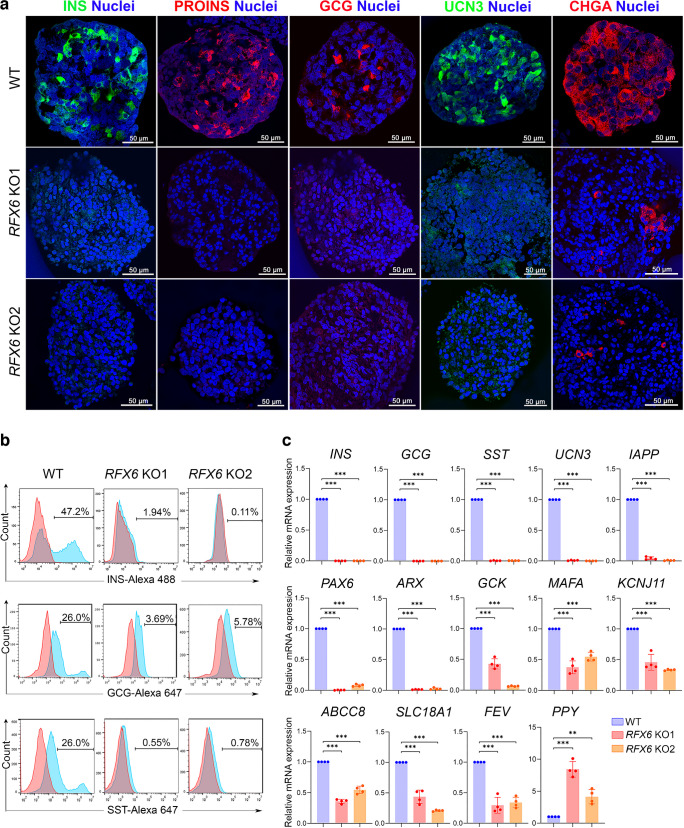
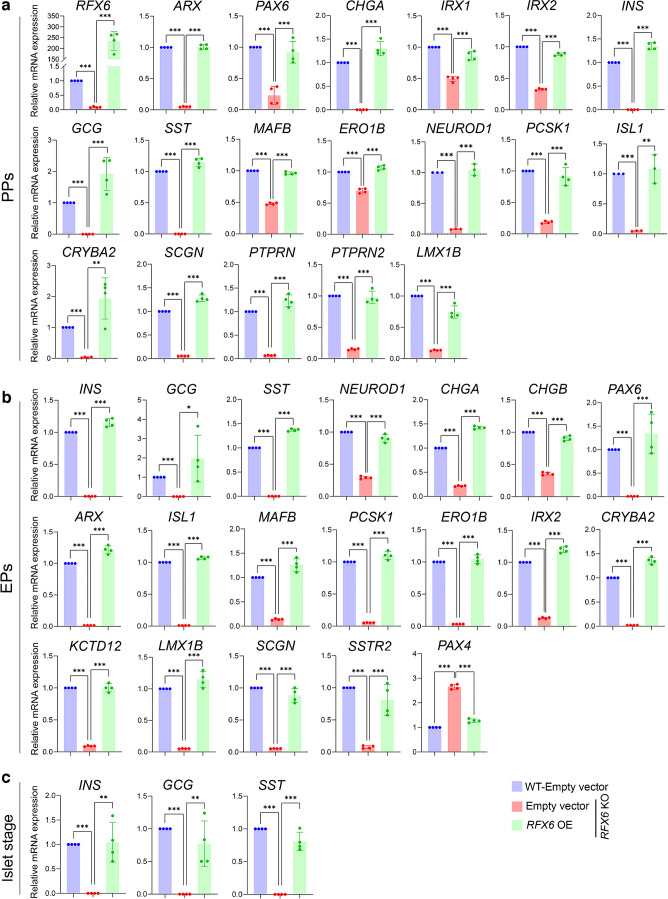
Results: RFX6 expression was detected in PDX1+ cells in the hESC-derived posterior foregut (PF). However, in the PPs, RFX6 did not co-localise with pancreatic and duodenal homeobox 1 (PDX1) or NK homeobox 1 (NKX6.1) but instead co-localised with neurogenin 3, NK2 homeobox 2 and islet hormones in the EPs and islets. Single-cell analysis revealed high RFX6 expression levels in endocrine clusters across various hESC-derived pancreatic differentiation stages. Upon differentiating iPSCs lacking RFX6 into pancreatic islets, a significant decrease in PDX1 expression at the PF stage was observed, although this did not affect PPs co-expressing PDX1 and NKX6.1. RNA-seq analysis showed the downregulation of essential genes involved in pancreatic endocrine differentiation, insulin secretion and ion transport due to RFX6 deficiency. Furthermore, RFX6 deficiency resulted in the formation of smaller islet organoids due to increased cellular apoptosis, linked to reduced catalase expression, implying a protective role for RFX6. Overexpression of RFX6 reversed defective phenotypes in RFX6-knockout PPs, EPs and islets.
Conclusions/interpretation: These findings suggest that pancreatic hypoplasia and reduced islet cell formation associated with RFX6 mutations are not due to alterations in PDX1+/NKX6.1+ PPs but instead result from cellular apoptosis and downregulation of pancreatic endocrine genes.
Data availability: RNA-seq datasets have been deposited in the Zenodo repository with accession link (DOI: https://doi.org/10.5281/zenodo.10656891 ).
Keywords: Diabetes; Endocrine specification; Islet organoids; Pancreatic hypoplasia; Pancreatic progenitors; Transcription factors.
© 2024. The Author(s).
Conflict of interest statement
Acknowledgements: We would like to thank N. R. Dunn (A*STAR, Singapore) for providing the HA-RFX6 tagged H9 hESC lines (RFX6HA/HA H9-hESCs). Furthermore, we thank the Genomic Core members at QBRI for their assistance with technical support in RNA-seq. Data availability: RNA-seq datasets have been deposited in the Zenodo repository with accession link (DOI: https://doi.org/10.5281/zenodo.10656891 ). Funding: Open Access funding provided by the Qatar National Library. This work was funded by grants from Qatar Biomedical Research Institute (QBRI) (Grant no. QBRI-HSCI Project 1). NA is a PhD student with a scholarship funded by QRDI (GSRA9-L-1-0511-22008). Authors’ relationships and activities: SH is a co-founder and shareholder of Sequantrix GmbH and has research funding from Novo Nordisk and Askbio. The authors declare that there are no other relationships or activities that might bias, or be perceived to bias, their work. Contribution statement: NA performed most of the experiments and analysed the data. AKE and BM performed experiments and analysed the data. SI and SH analysed the sequencing data. EMA conceived and designed the study, supervised the project, analysed and interpreted the data, and wrote the manuscript. All authors critically reviewed the article and approved the final version of the manuscript. EMA is the guarantor of this work.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
