

Myeloid PTEN loss affects the therapeutic response by promoting stress granule assembly and impairing phagocytosis by macrophages in breast cancer
- PMID: 39080255
- PMCID: PMC11289284
- DOI: 10.1038/s41420-024-02094-0
Myeloid PTEN loss affects the therapeutic response by promoting stress granule assembly and impairing phagocytosis by macrophages in breast cancer
Abstract
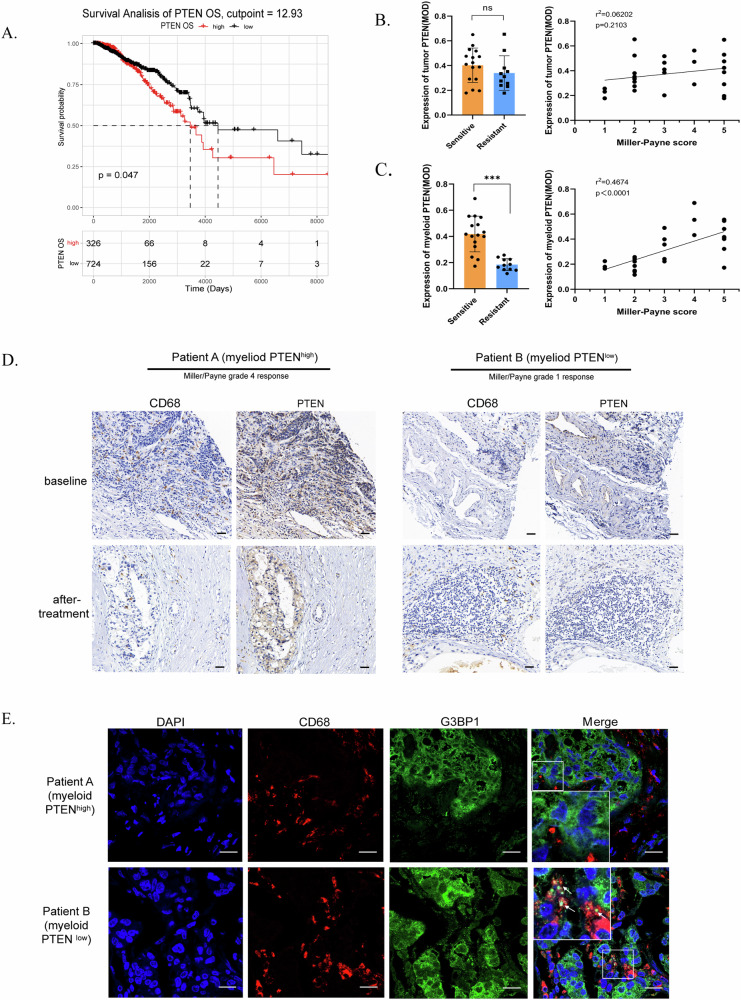
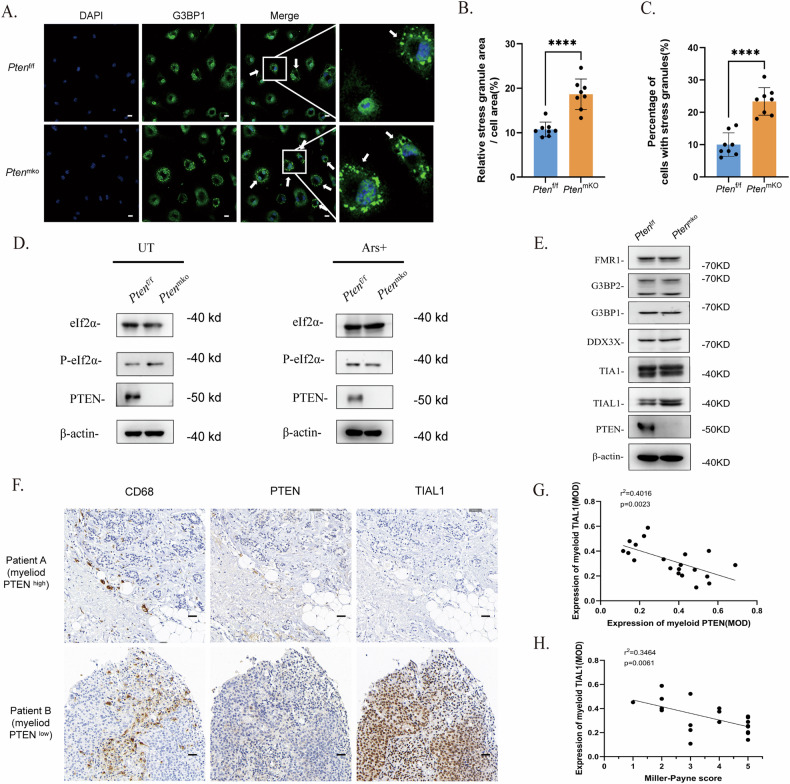
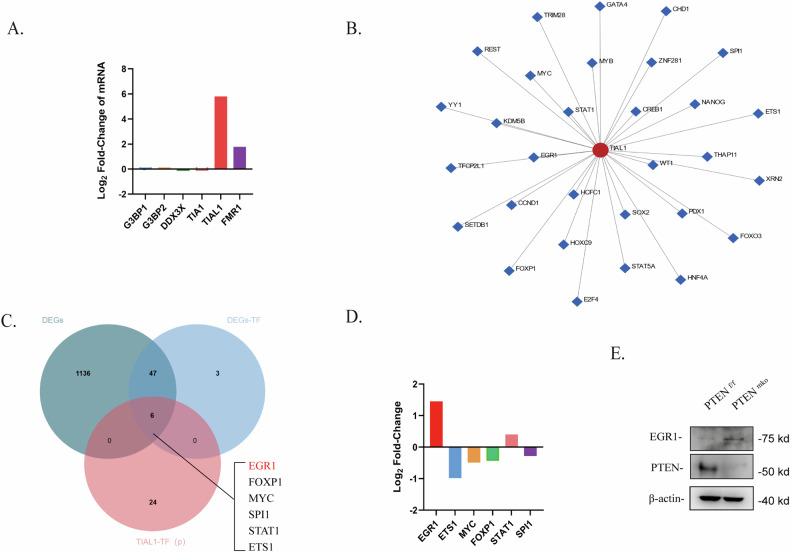
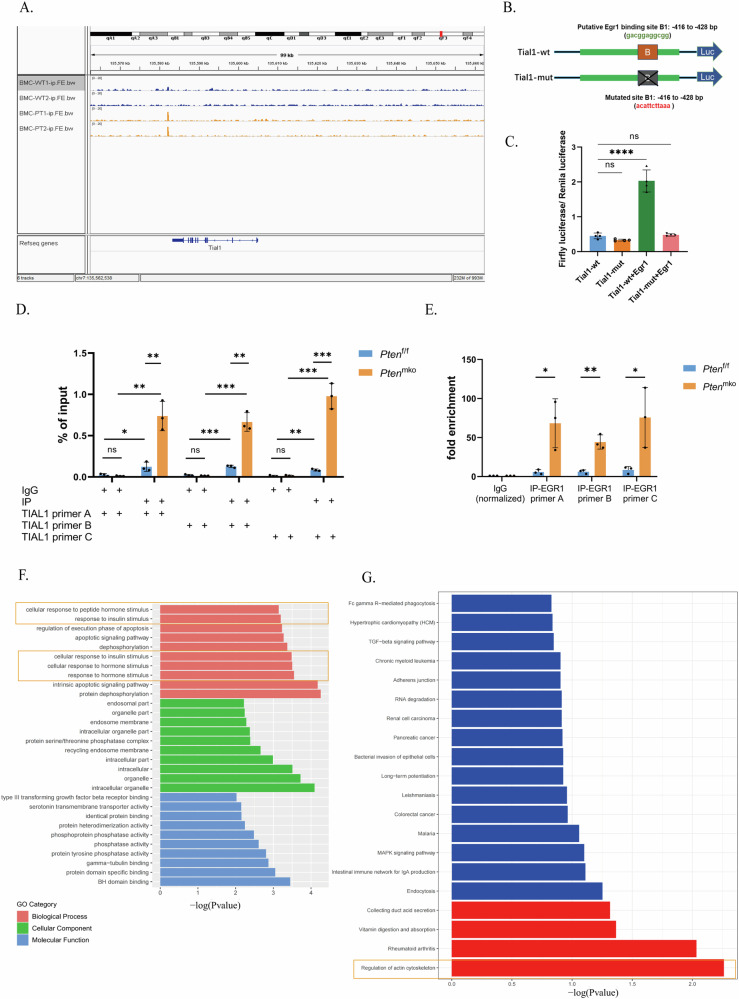
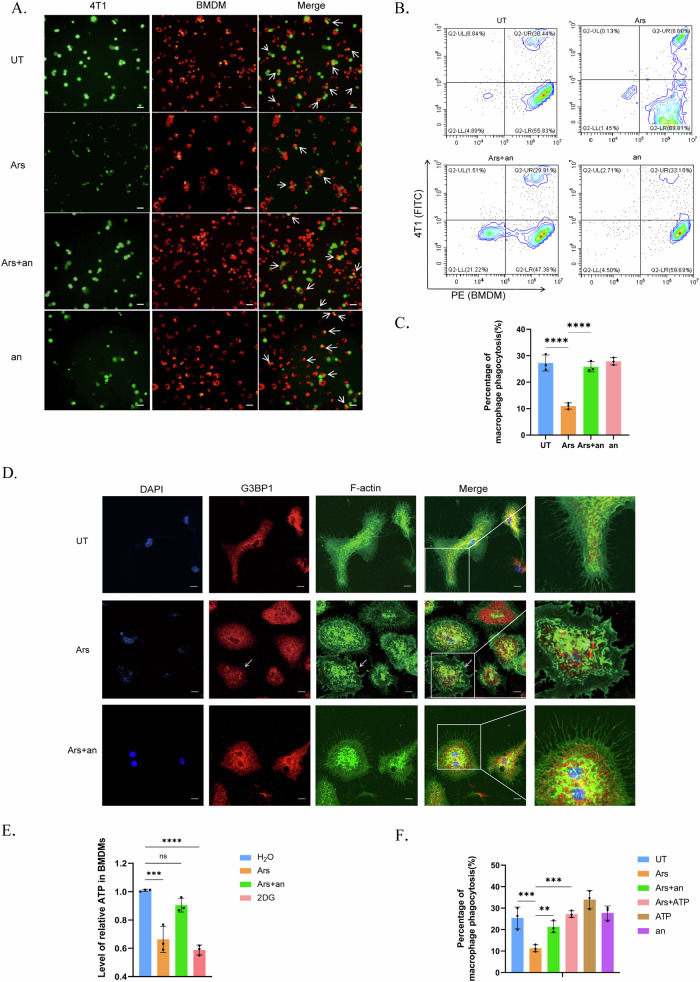
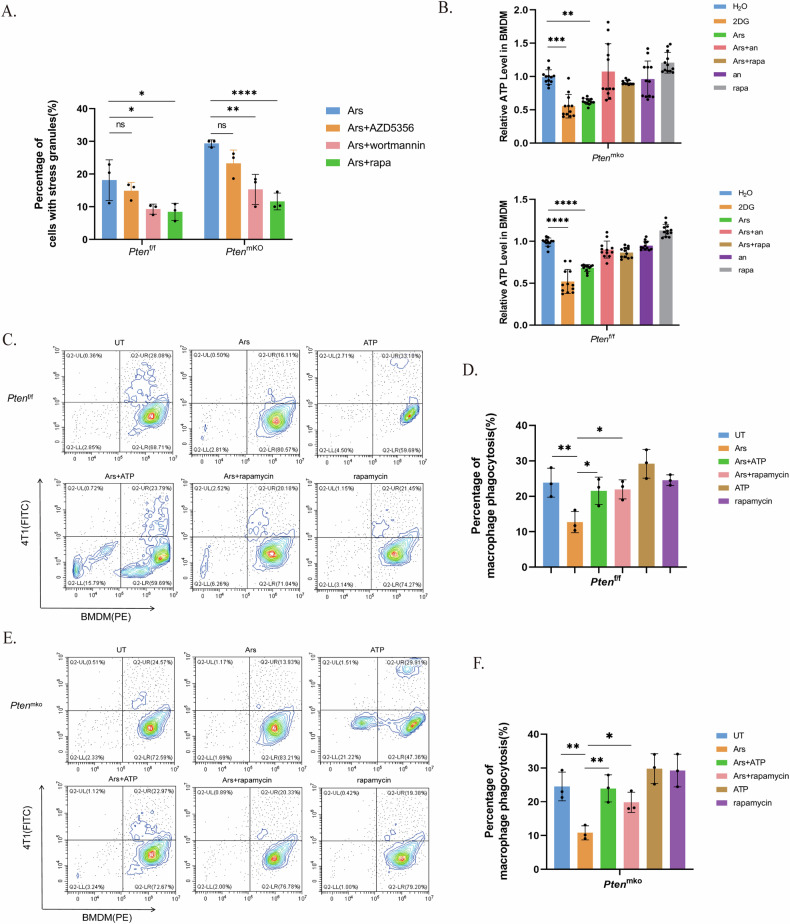
Breast cancer (BRCA) has become the most common type of cancer in women. Improving the therapeutic response remains a challenge. Phosphatase and tensin homologue deleted on chromosome 10 (PTEN) is a classic tumour suppressor with emerging new functions discovered in recent years, and myeloid PTEN loss has been reported to impair antitumour immunity. In this study, we revealed a novel mechanism by which myeloid PTEN potentially affects antitumour immunity in BRCA. We detected accelerated stress granule (SG) assembly under oxidative stress in PTEN-deficient bone marrow-derived macrophages (BMDMs) through the EGR1-promoted upregulation of TIAL1 transcription. PI3K/AKT/mTOR (PAM) pathway activation also promoted SG formation. ATP consumption during SG assembly in BMDMs impaired the phagocytic ability of 4T1 cells, potentially contributing to the disruption of antitumour immunity. In a BRCA neoadjuvant cohort, we observed a poorer response in myeloid PTENlow patients with G3BP1 aggregating as SGs in CD68+ cells, a finding that was consistent with the observation in our study that PTEN-deficient macrophages tended to more readily assemble SGs with impaired phagocytosis. Our results revealed the unconventional impact of SGs on BMDMs and might provide new perspectives on drug resistance and therapeutic strategies for the treatment of BRCA patients.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Myeloid PTEN promotes chemotherapy-induced NLRP3-inflammasome activation and antitumour immunity.Nat Cell Biol. 2020 Jun;22(6):716-727. doi: 10.1038/s41556-020-0510-3. Epub 2020 May 4. Nat Cell Biol. 2020. PMID: 32367047
-
Stress granule homeostasis is modulated by TRIM21-mediated ubiquitination of G3BP1 and autophagy-dependent elimination of stress granules.Autophagy. 2023 Jul;19(7):1934-1951. doi: 10.1080/15548627.2022.2164427. Epub 2023 Jan 24. Autophagy. 2023. PMID: 36692217 Free PMC article.
-
The inositol 3-phosphatase PTEN negatively regulates Fc gamma receptor signaling, but supports Toll-like receptor 4 signaling in murine peritoneal macrophages.J Immunol. 2004 Apr 15;172(8):4851-7. doi: 10.4049/jimmunol.172.8.4851. J Immunol. 2004. PMID: 15067063
-
The INPP4B paradox: Like PTEN, but different.Adv Biol Regul. 2021 Dec;82:100817. doi: 10.1016/j.jbior.2021.100817. Epub 2021 Jun 16. Adv Biol Regul. 2021. PMID: 34216856 Review.
-
New insights into Notch1 regulation of the PI3K-AKT-mTOR1 signaling axis: targeted therapy of γ-secretase inhibitor resistant T-cell acute lymphoblastic leukemia.Cell Signal. 2014 Jan;26(1):149-61. doi: 10.1016/j.cellsig.2013.09.021. Epub 2013 Oct 16. Cell Signal. 2014. PMID: 24140475 Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
