

Coupling sensor to enzyme in the voltage sensing phosphatase
- PMID: 39080263
- PMCID: PMC11289409
- DOI: 10.1038/s41467-024-50319-8
Coupling sensor to enzyme in the voltage sensing phosphatase
Abstract
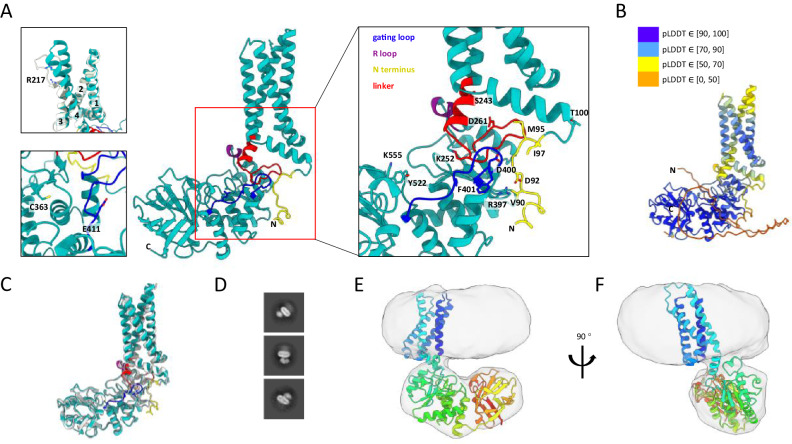
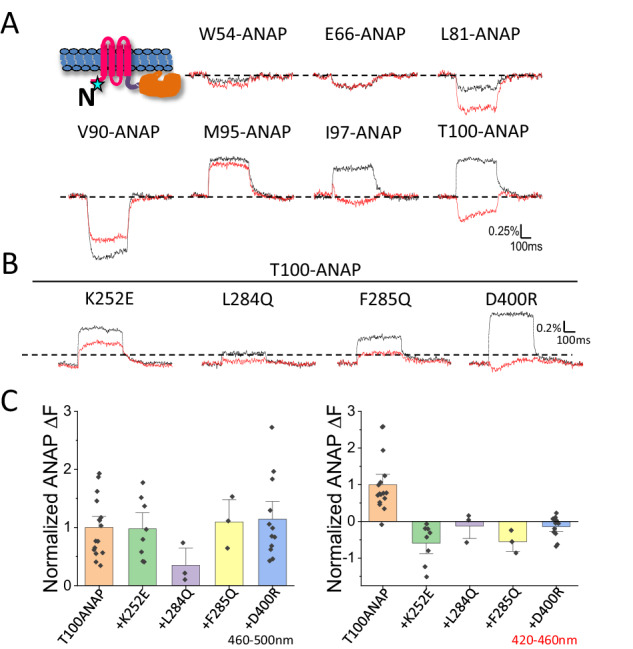
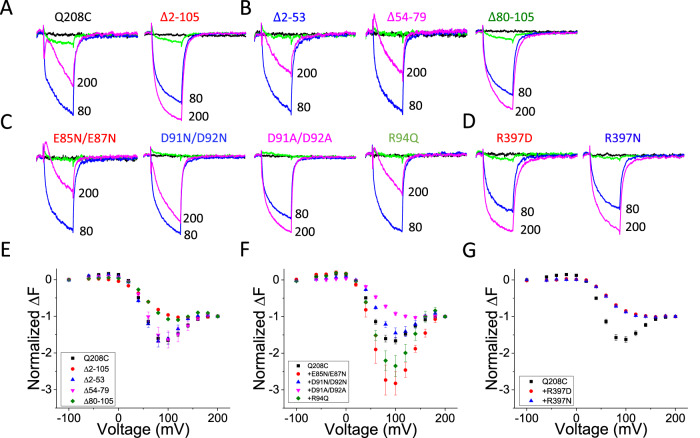
Voltage-sensing phosphatases (VSPs) dephosphorylate phosphoinositide (PIP) signaling lipids in response to membrane depolarization. VSPs possess an S4-containing voltage sensor domain (VSD), resembling that of voltage-gated cation channels, and a lipid phosphatase domain (PD). The mechanism by which voltage turns on enzyme activity is unclear. Structural analysis and modeling suggest several sites of VSD-PD interaction that could couple voltage sensing to catalysis. Voltage clamp fluorometry reveals voltage-driven rearrangements in three sites implicated earlier in enzyme activation-the VSD-PD linker, gating loop and R loop-as well as the N-terminal domain, which has not yet been explored. N-terminus mutations perturb both rearrangements in the other segments and enzyme activity. Our results provide a model for a dynamic assembly by which S4 controls the catalytic site.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interest.
Figures





Similar articles
-
Interaction between S4 and the phosphatase domain mediates electrochemical coupling in voltage-sensing phosphatase (VSP).Proc Natl Acad Sci U S A. 2022 Jun 28;119(26):e2200364119. doi: 10.1073/pnas.2200364119. Epub 2022 Jun 21. Proc Natl Acad Sci U S A. 2022. PMID: 35733115 Free PMC article.
-
Electrochemical coupling in the voltage-dependent phosphatase Ci-VSP.Nat Chem Biol. 2010 May;6(5):369-75. doi: 10.1038/nchembio.349. Epub 2010 Apr 4. Nat Chem Biol. 2010. PMID: 20364128 Free PMC article.
-
The hydrophobic nature of a novel membrane interface regulates the enzyme activity of a voltage-sensing phosphatase.Elife. 2018 Nov 28;7:e41653. doi: 10.7554/eLife.41653. Elife. 2018. PMID: 30484774 Free PMC article.
-
Voltage-Sensing Phosphatases: Biophysics, Physiology, and Molecular Engineering.Physiol Rev. 2018 Oct 1;98(4):2097-2131. doi: 10.1152/physrev.00056.2017. Physiol Rev. 2018. PMID: 30067160 Review.
-
Dynamic structural rearrangements and functional regulation of voltage-sensing phosphatase.J Physiol. 2019 Jan;597(1):29-40. doi: 10.1113/JP274113. Epub 2018 Nov 22. J Physiol. 2019. PMID: 30311949 Free PMC article. Review.
Cited by
-
Noncoding RNA-encoded peptides in cancer: biological functions, posttranslational modifications and therapeutic potential.J Hematol Oncol. 2025 Feb 19;18(1):20. doi: 10.1186/s13045-025-01671-9. J Hematol Oncol. 2025. PMID: 39972384 Free PMC article. Review.
-
Nonsubstrate PI(4,5)P2 interacts with the interdomain linker to control electrochemical coupling in voltage-sensing phosphatase (VSP).Proc Natl Acad Sci U S A. 2025 Aug 5;122(31):e2500651122. doi: 10.1073/pnas.2500651122. Epub 2025 Jul 29. Proc Natl Acad Sci U S A. 2025. PMID: 40729387
References
MeSH terms
Substances
Grants and funding
- R01GM117051/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- GM145869/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- R01 GM145869/GM/NIGMS NIH HHS/United States
- R01 GM117051/GM/NIGMS NIH HHS/United States
- F32 NS101816/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
