

Enhancers associated with unstable RNAs are rare in plants
- PMID: 39080503
- PMCID: PMC11335568
- DOI: 10.1038/s41477-024-01741-9
Enhancers associated with unstable RNAs are rare in plants
Abstract
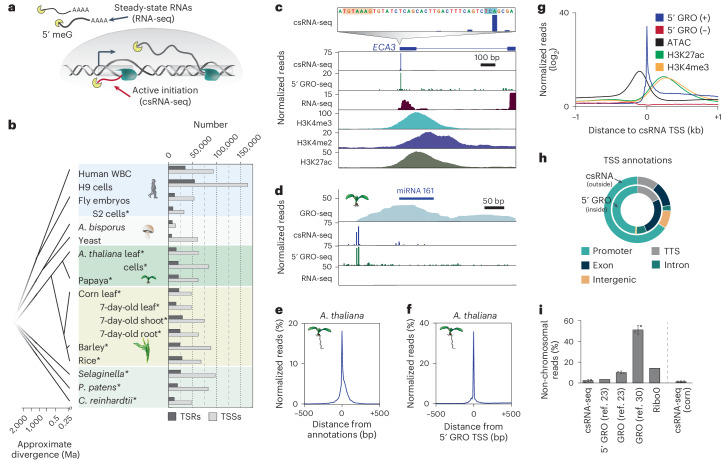
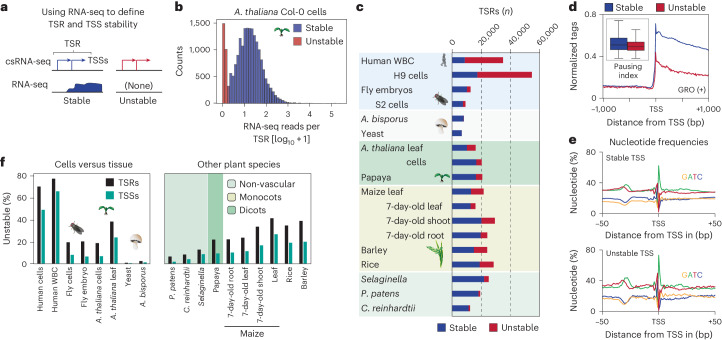
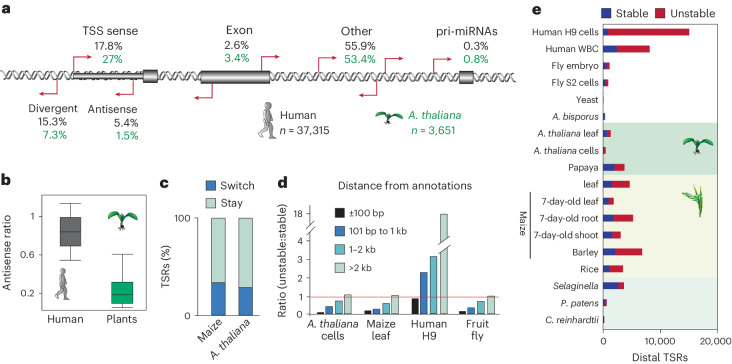
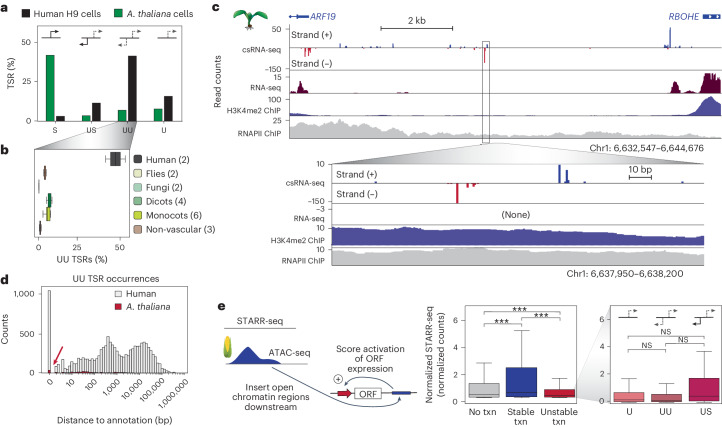
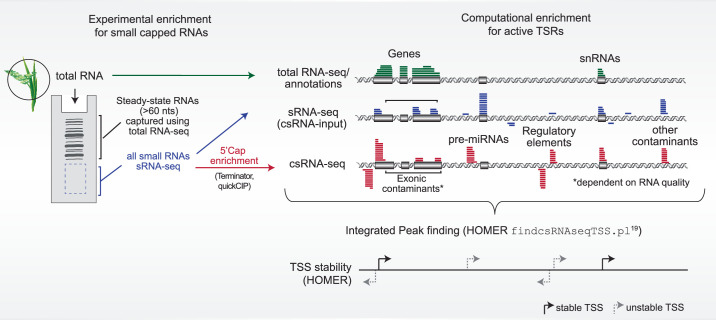
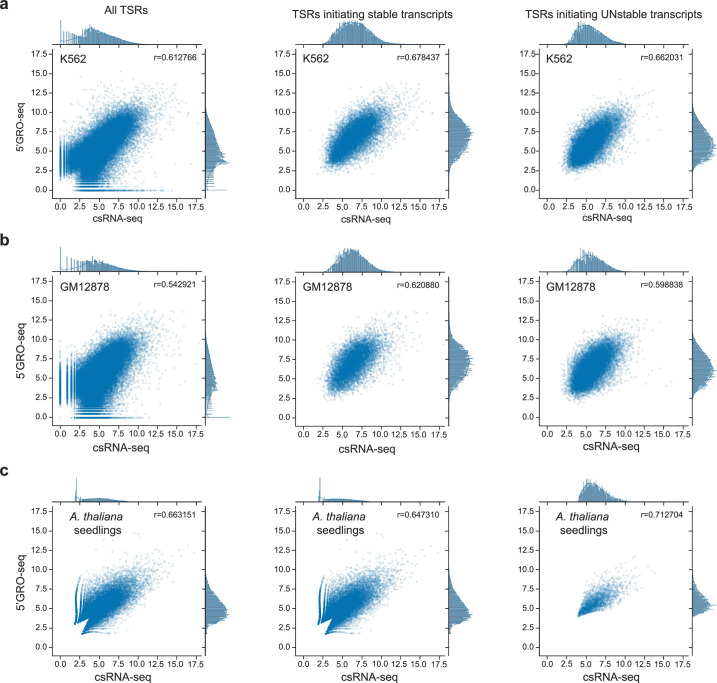
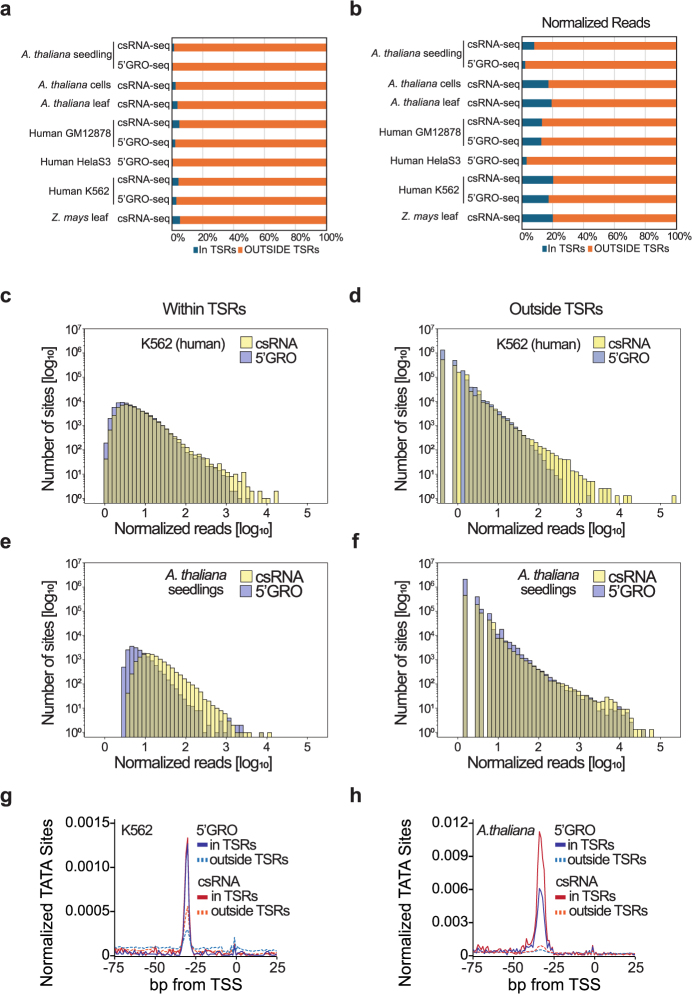
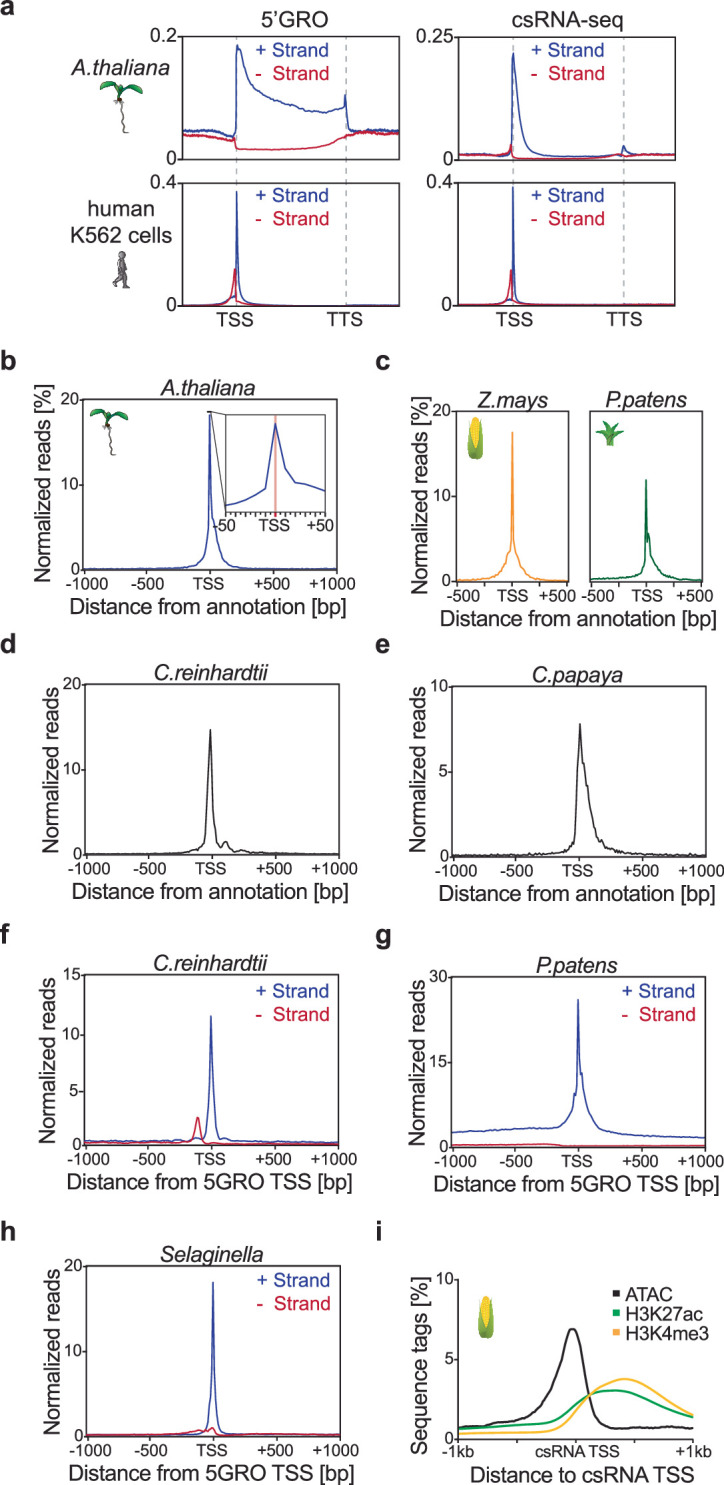
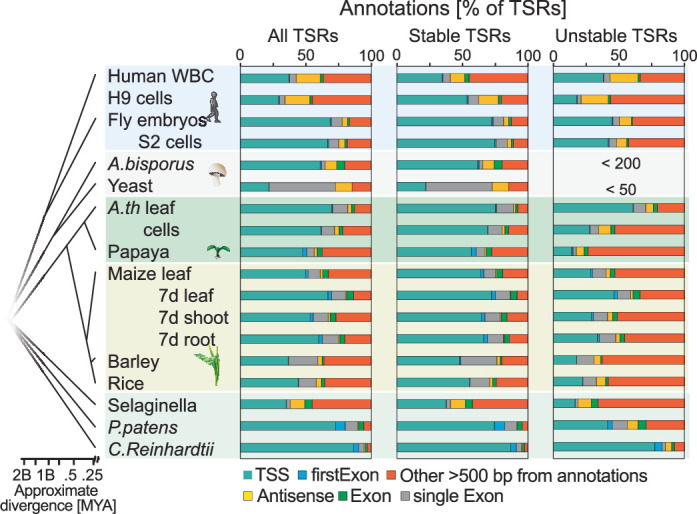
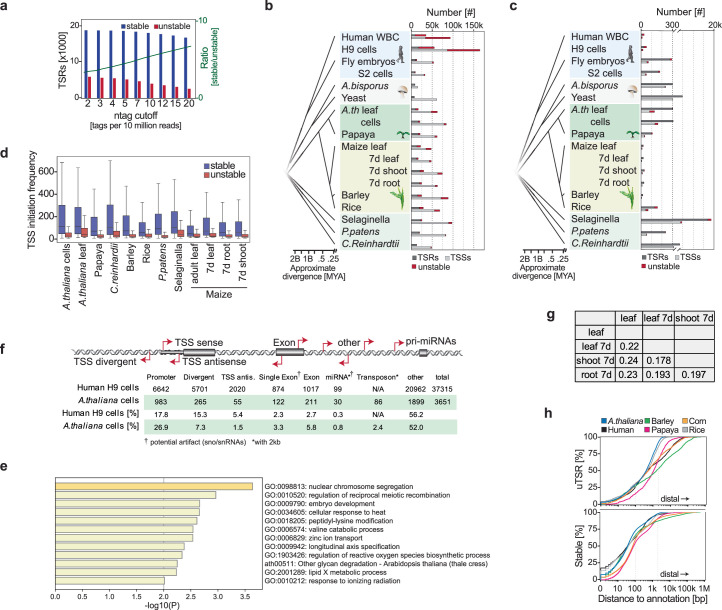
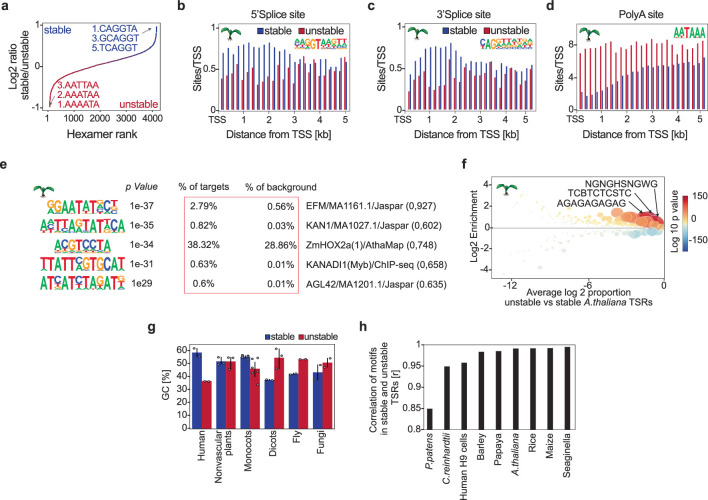
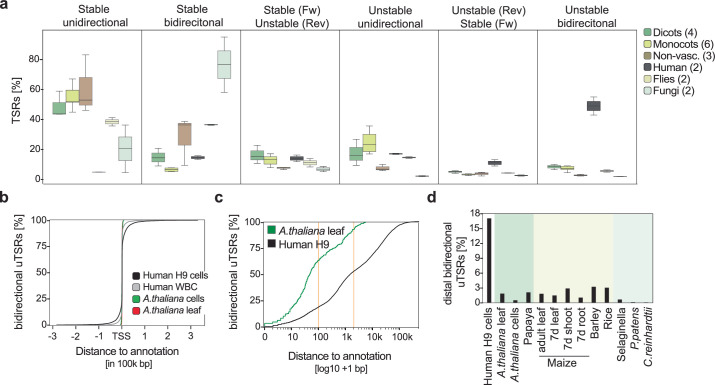
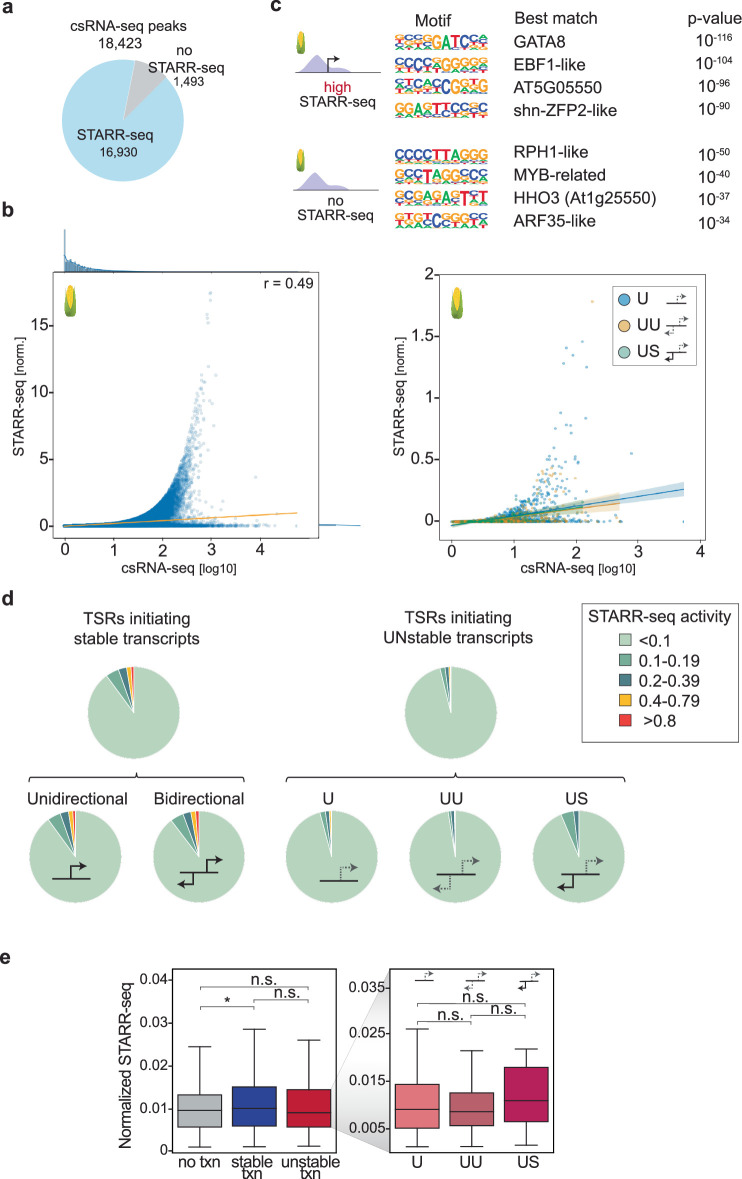
Unstable transcripts have emerged as markers of active enhancers in vertebrates and shown to be involved in many cellular processes and medical disorders. However, their prevalence and role in plants is largely unexplored. Here, we comprehensively captured all actively initiating (nascent) transcripts across diverse crops and other plants using capped small (cs)RNA sequencing. We discovered that unstable transcripts are rare in plants, unlike in vertebrates, and when present, often originate from promoters. In addition, many 'distal' elements in plants initiate tissue-specific stable transcripts and are likely bona fide promoters of as-yet-unannotated genes or non-coding RNAs, cautioning against using reference genome annotations to infer putative enhancer sites. To investigate enhancer function, we integrated data from self-transcribing active regulatory region (STARR) sequencing. We found that annotated promoters and other regions that initiate stable transcripts, but not those marked by unstable or bidirectional unstable transcripts, showed stronger enhancer activity in this assay. Our findings underscore the blurred line between promoters and enhancers and suggest that cis-regulatory elements can encompass diverse structures and mechanisms in eukaryotes, including humans.
© 2024. This is a U.S. Government work and not under copyright protection in the US; foreign copyright protection may apply.
Conflict of interest statement
The authors declare no competing interests.
Figures
Update of
-
Enhancers associated with unstable RNAs are rare in plants.bioRxiv [Preprint]. 2023 Sep 25:2023.09.25.559415. doi: 10.1101/2023.09.25.559415. bioRxiv. 2023. Update in: Nat Plants. 2024 Aug;10(8):1246-1257. doi: 10.1038/s41477-024-01741-9. PMID: 37808859 Free PMC article. Updated. Preprint.
Similar articles
-
Enhancers associated with unstable RNAs are rare in plants.bioRxiv [Preprint]. 2023 Sep 25:2023.09.25.559415. doi: 10.1101/2023.09.25.559415. bioRxiv. 2023. Update in: Nat Plants. 2024 Aug;10(8):1246-1257. doi: 10.1038/s41477-024-01741-9. PMID: 37808859 Free PMC article. Updated. Preprint.
-
Compatibility rules of human enhancer and promoter sequences.Nature. 2022 Jul;607(7917):176-184. doi: 10.1038/s41586-022-04877-w. Epub 2022 May 20. Nature. 2022. PMID: 35594906 Free PMC article.
-
Single-cell nascent RNA sequencing unveils coordinated global transcription.Nature. 2024 Jul;631(8019):216-223. doi: 10.1038/s41586-024-07517-7. Epub 2024 Jun 5. Nature. 2024. PMID: 38839954 Free PMC article.
-
Enhancer RNAs: a missing regulatory layer in gene transcription.Sci China Life Sci. 2019 Jul;62(7):905-912. doi: 10.1007/s11427-017-9370-9. Epub 2018 Dec 26. Sci China Life Sci. 2019. PMID: 30593613 Review.
-
Towards genome-wide prediction and characterization of enhancers in plants.Biochim Biophys Acta Gene Regul Mech. 2017 Jan;1860(1):131-139. doi: 10.1016/j.bbagrm.2016.06.006. Epub 2016 Jun 16. Biochim Biophys Acta Gene Regul Mech. 2017. PMID: 27321818 Review.
Cited by
-
Integrated expression analysis to elucidate the role of miR394 during flowering in Arabidopsis thaliana.Plant Cell Rep. 2025 Jul 2;44(7):167. doi: 10.1007/s00299-025-03541-7. Plant Cell Rep. 2025. PMID: 40601081
-
Genome organization and botanical diversity.Plant Cell. 2024 May 1;36(5):1186-1204. doi: 10.1093/plcell/koae045. Plant Cell. 2024. PMID: 38382084 Free PMC article. Review.
-
MaizeCODE reveals bi-directionally expressed enhancers that harbor molecular signatures of maize domestication.Nat Commun. 2024 Dec 30;15(1):10854. doi: 10.1038/s41467-024-55195-w. Nat Commun. 2024. PMID: 39738013 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
