

Lactylation of histone by BRD4 regulates astrocyte polarization after experimental subarachnoid hemorrhage
- PMID: 39080649
- PMCID: PMC11290164
- DOI: 10.1186/s12974-024-03185-6
Lactylation of histone by BRD4 regulates astrocyte polarization after experimental subarachnoid hemorrhage
Abstract
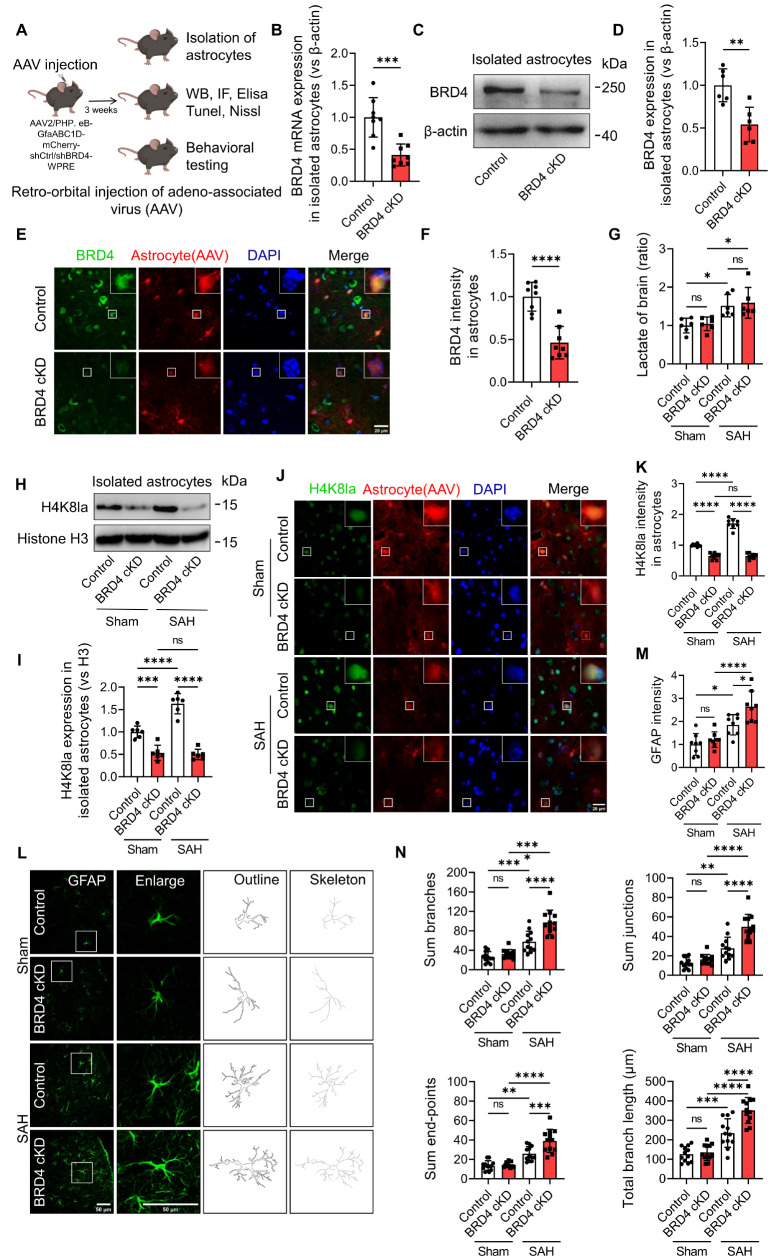
Under subarachnoid hemorrhage (SAH) conditions, astrocytes undergo a marked intensification of glycolytic activity, resulting in the generation of substantial amounts of lactate to maintain the energy demand for neurons and other brain cells. Lactate has garnered increasing attention in recent years because of its emerging role in critical biological processes such as inflammation regulation and neuroprotection, particularly through its histone lactylation. Bromodomain-containing protein 4 (BRD4) plays a crucial role in maintaining neural development and promoting memory formation in the central nervous system. Nonetheless, the function and regulatory mechanism of BRD4 and histone lactylation in astrocytes following SAH remain elusive. Our findings indicate that BRD4, a crucial epigenetic regulator, plays a definitive role in histone lactylation. Both in vitro and in vivo, these results demonstrated that targeted silencing of BRD4 in astrocytes can significantly reduce H4K8la lactylation, thereby aggravating the A1 polarization of astrocytes and ultimately affecting the recovery of neural function and prognosis in mice after SAH. In summary, BRD4 plays a pivotal role in modulating astrocyte polarization following SAH via histone lactylation. Targeting this mechanism might offer an efficient therapeutic strategy for SAH.
Keywords: Astrocytic polarization; Bromodomain-containing protein 4; Histone lactylation; Subarachnoid hemorrhage.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures







References
-
- Etminan N, Chang HS, Hackenberg K, et al. Worldwide Incidence of Aneurysmal Subarachnoid Hemorrhage according to Region, Time Period, blood pressure, and Smoking Prevalence in the Population: a systematic review and Meta-analysis. JAMA Neurol. 2019;76(5):588–97. 10.1001/jamaneurol.2019.0006 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- 2023NSFSC1559/Sichuan Province Science and Technology Support Program
- 2022NSFSC1421/Sichuan Province Science and Technology Support Program
- 2022YFS0615/Sichuan Province Science and Technology Support Program
- 2023NSFSC0028/Sichuan Province Science and Technology Support Program
- 2023YFH0069/Sichuan Province Science and Technology Support Program
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
