

MicroRNA-19b exacerbates systemic sclerosis through promoting Th9 cells
- PMID: 39083380
- PMCID: PMC11440512
- DOI: 10.1016/j.celrep.2024.114565
MicroRNA-19b exacerbates systemic sclerosis through promoting Th9 cells
Erratum in
-
MicroRNA-19b exacerbates systemic sclerosis through promoting Th9 cells.Cell Rep. 2025 Mar 25;44(3):115447. doi: 10.1016/j.celrep.2025.115447. Epub 2025 Mar 5. Cell Rep. 2025. PMID: 40048430 No abstract available.
Abstract
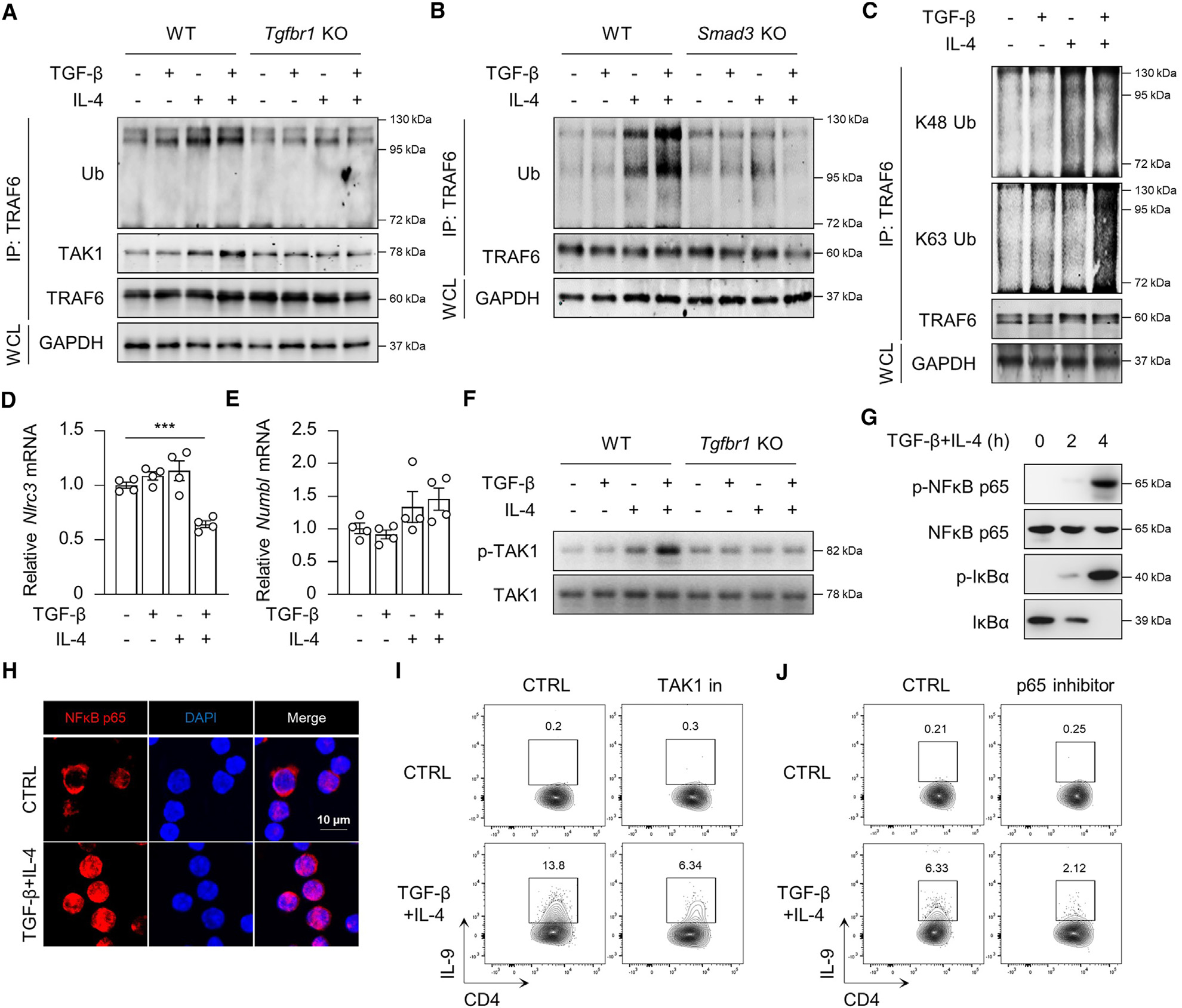
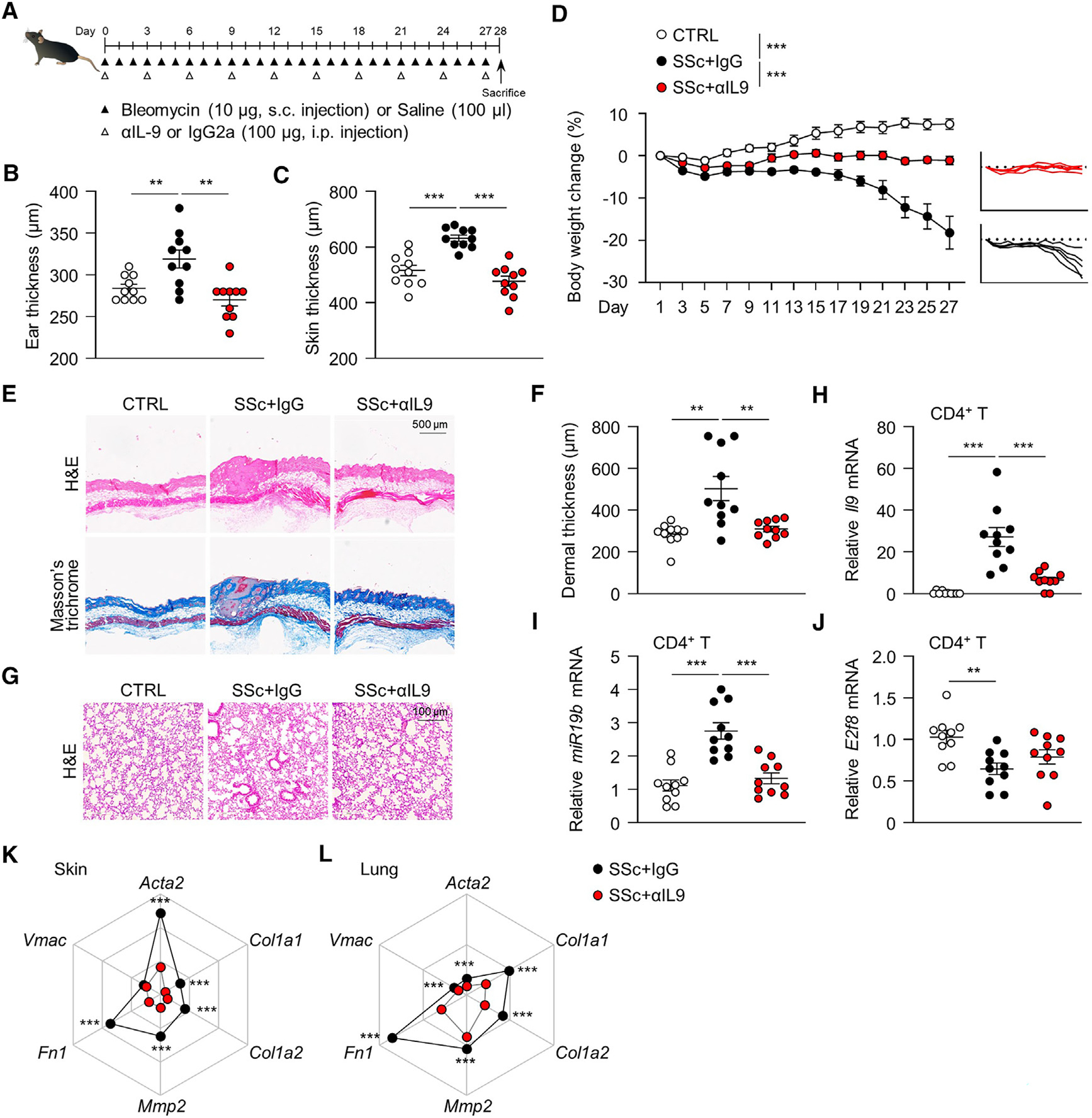
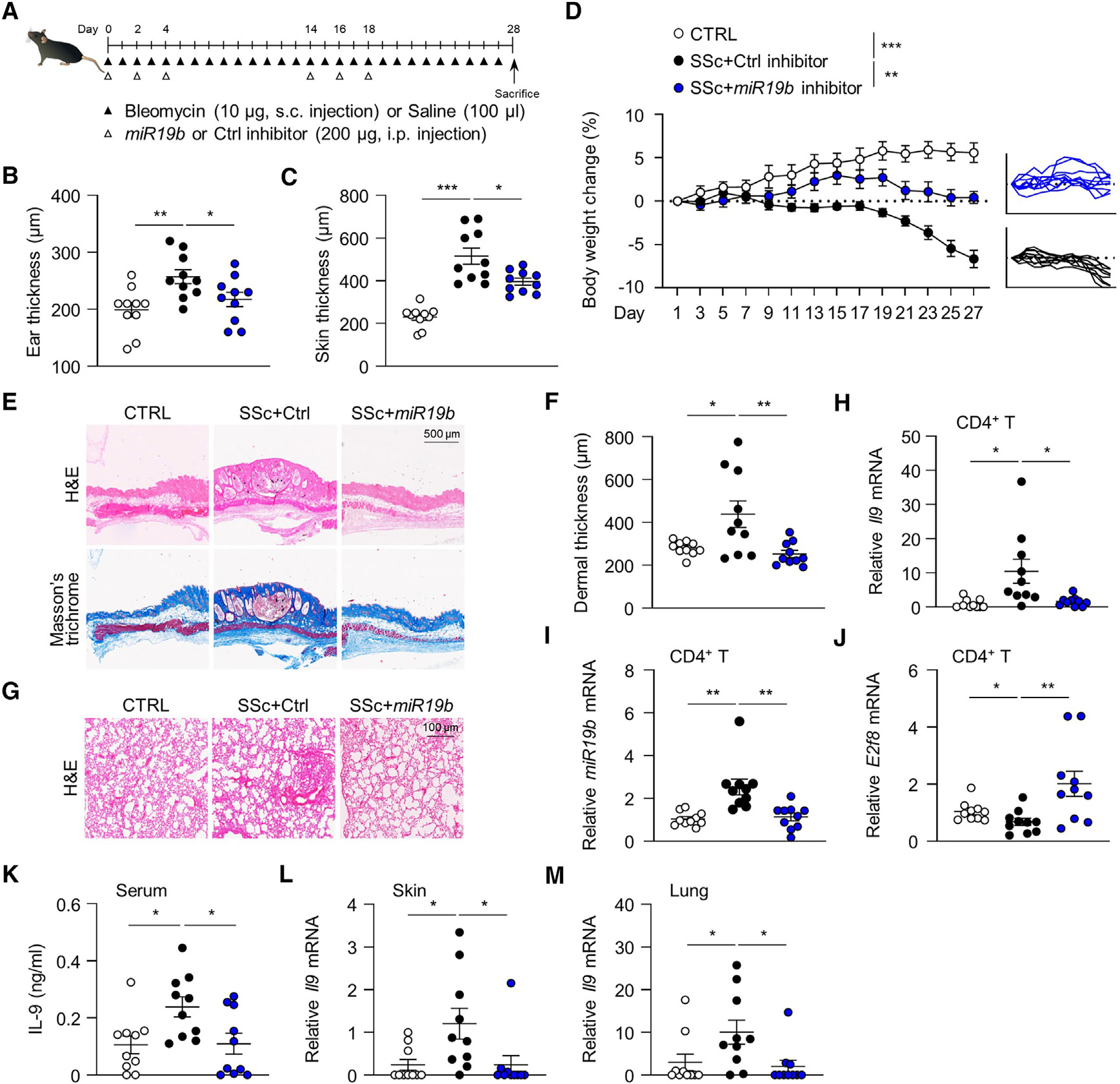
Systemic sclerosis (SSc) is a chronic autoimmune disease characterized by fibrosis of the skin and multiple vital organs, but the immunological pathogenesis of SSc remains unclear. We show here that miR-19b promotes Th9 cells that exacerbate SSc. Specifically, miR-19b and interleukin (IL)-9 increase in CD4+ T cells in experimental SSc in mice induced with bleomycin. Inhibiting miR-19b reduces Th9 cells and ameliorates the disease. Mechanistically, transforming growth factor beta (TGF-β) plus IL-4 activates pSmad3-Ser213 and TRAF6-K63 ubiquitination by suppressing NLRC3. Activated TRAF6 sequentially promotes TGF-β-activated kinase 1 (TAK1) and nuclear factor κB (NF-κB) p65 phosphorylation, leading to the upregulation of miR-19b. Notably, miR-19b activated Il9 gene expression by directly suppressing atypical E2F family member E2f8. In patients with SSc, higher levels of IL9 and MIR-19B correlate with worse disease progression. Our findings reveal miR-19b as a key factor in Th9 cell-mediated SSc pathogenesis and should have clinical implications for patients with SSc.
Keywords: CP: Immunology; E2f8; NF-κB p65; TAK1; TGF-β signaling; TRAF6; Th9; miR-19b; patients with systemic sclerosis.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
