

Immediate myeloid depot for SARS-CoV-2 in the human lung
- PMID: 39083602
- PMCID: PMC11290487
- DOI: 10.1126/sciadv.adm8836
Immediate myeloid depot for SARS-CoV-2 in the human lung
Abstract
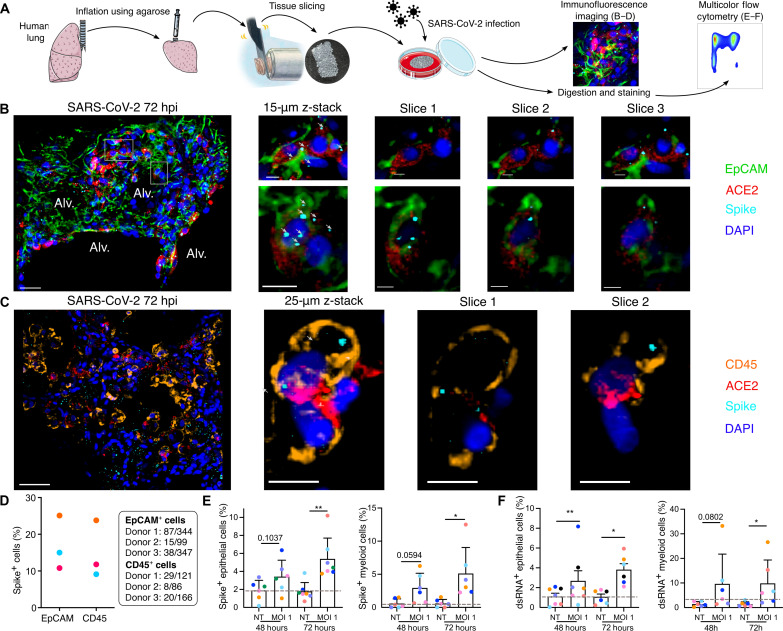
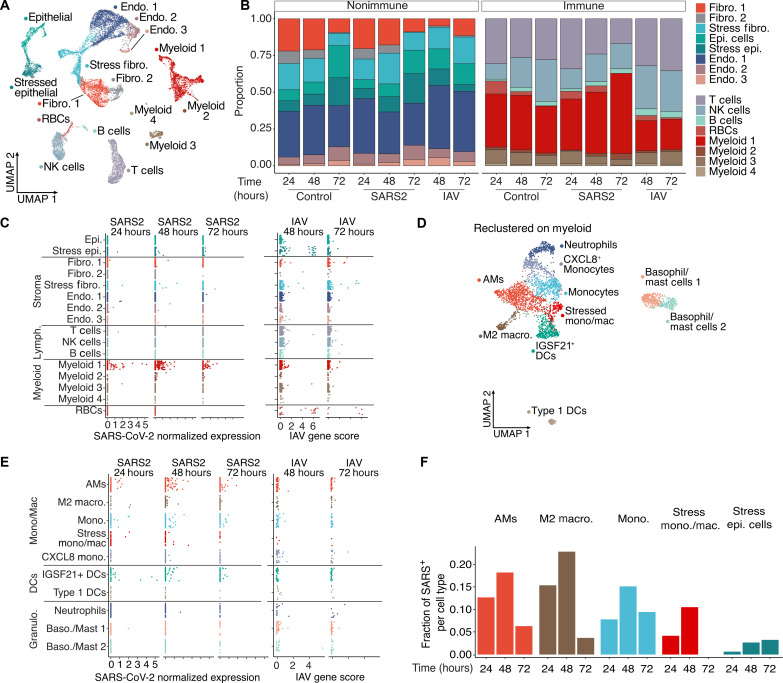
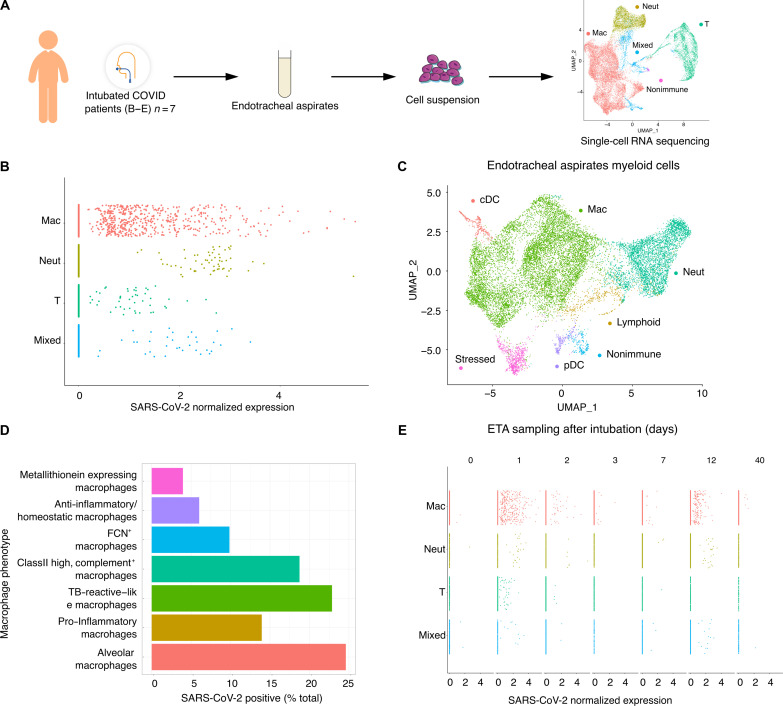
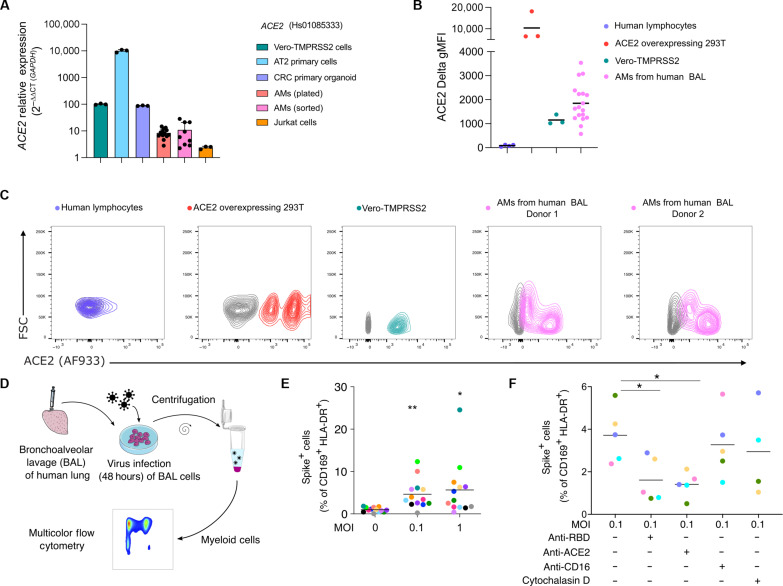
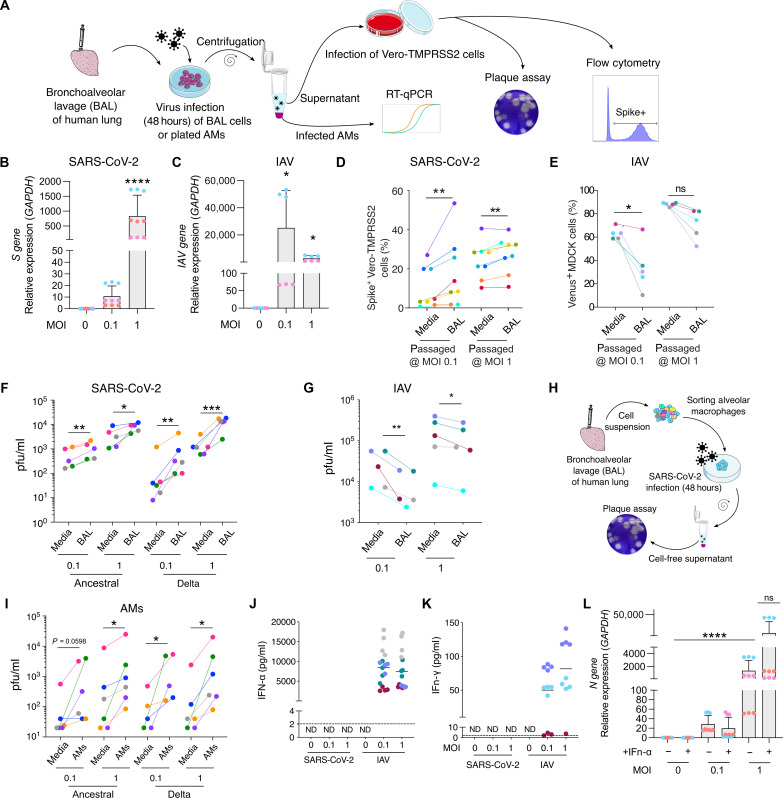
In the pathogenesis of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection, epithelial populations in the distal lung expressing Angiotensin-converting enzyme 2 (ACE2) are infrequent, and therefore, the model of viral expansion and immune cell engagement remains incompletely understood. Using human lungs to investigate early host-viral pathogenesis, we found that SARS-CoV-2 had a rapid and specific tropism for myeloid populations. Human alveolar macrophages (AMs) reliably expressed ACE2 allowing both spike-ACE2-dependent viral entry and infection. In contrast to Influenza A virus, SARS-CoV-2 infection of AMs was productive, amplifying viral titers. While AMs generated new viruses, the interferon responses to SARS-CoV-2 were muted, hiding the viral dissemination from specific antiviral immune responses. The reliable and veiled viral depot in myeloid cells in the very early phases of SARS-CoV-2 infection of human lungs enables viral expansion in the distal lung and potentially licenses subsequent immune pathologies.
Figures
Update of
-
Immediate myeloid depot for SARS-CoV-2 in the human lung.bioRxiv [Preprint]. 2022 May 11:2022.04.28.489942. doi: 10.1101/2022.04.28.489942. bioRxiv. 2022. Update in: Sci Adv. 2024 Aug 2;10(31):eadm8836. doi: 10.1126/sciadv.adm8836. PMID: 35592107 Free PMC article. Updated. Preprint.
-
Immediate myeloid depot for SARS-CoV-2 in the human lung.Res Sq [Preprint]. 2022 May 17:rs.3.rs-1639631. doi: 10.21203/rs.3.rs-1639631/v1. Res Sq. 2022. Update in: Sci Adv. 2024 Aug 2;10(31):eadm8836. doi: 10.1126/sciadv.adm8836. PMID: 35611333 Free PMC article. Updated. Preprint.
References
-
- Lamers M. M., Haagmans B. L., SARS-CoV-2 pathogenesis. Nat. Rev. Microbiol. 20, 270–284 (2022). - PubMed
-
- Combes A. J., Courau T., Kuhn N. F., Hu K. H., Ray A., Chen W. S., Chew N. W., Cleary S. J., Kushnoor D., Reeder G. C., Shen A., Tsui J., Hiam-Galvez K. J., Munoz-Sandoval P., Zhu W. S., Lee D. S., Sun Y., You R., Magnen M., Rodriguez L., Im K. W., Serwas N. K., Leligdowicz A., Zamecnik C. R., Loudermilk R. P., Wilson M. R., Ye C. J., Fragiadakis G. K., Looney M. R., Chan V., Ward A., Carrillo S., Consortium U. C., Matthay M., Erle D. J., Woodruff P. G., Langelier C., Kangelaris K., Hendrickson C. M., Calfee C., Rao A. A., Krummel M. F., Global absence and targeting of protective immune states in severe COVID-19. Nature 591, 124–130 (2021). - PMC - PubMed
-
- Mathew D., Giles J. R., Baxter A. E., Oldridge D. A., Greenplate A. R., Wu J. E., Alanio C., Kuri-Cervantes L., Pampena M. B., D’Andrea K., Manne S., Chen Z., Huang Y. J., Reilly J. P., Weisman A. R., Ittner C. A. G., Kuthuru O., Dougherty J., Nzingha K., Han N., Kim J., Pattekar A., Goodwin E. C., Anderson E. M., Weirick M. E., Gouma S., Arevalo C. P., Bolton M. J., Chen F., Lacey S. F., Ramage H., Cherry S., Hensley S. E., Apostolidis S. A., Huang A. C., Vella L. A.; UPenn COVID Processing Unit, Betts M. R., Meyer N. J., Wherry E. J., Deep immune profiling of COVID-19 patients reveals distinct immunotypes with therapeutic implications. Science 369, eabc8511 (2020). - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
