

Eg5 UFMylation promotes spindle organization during mitosis
- PMID: 39085203
- PMCID: PMC11291904
- DOI: 10.1038/s41419-024-06934-w
Eg5 UFMylation promotes spindle organization during mitosis
Abstract
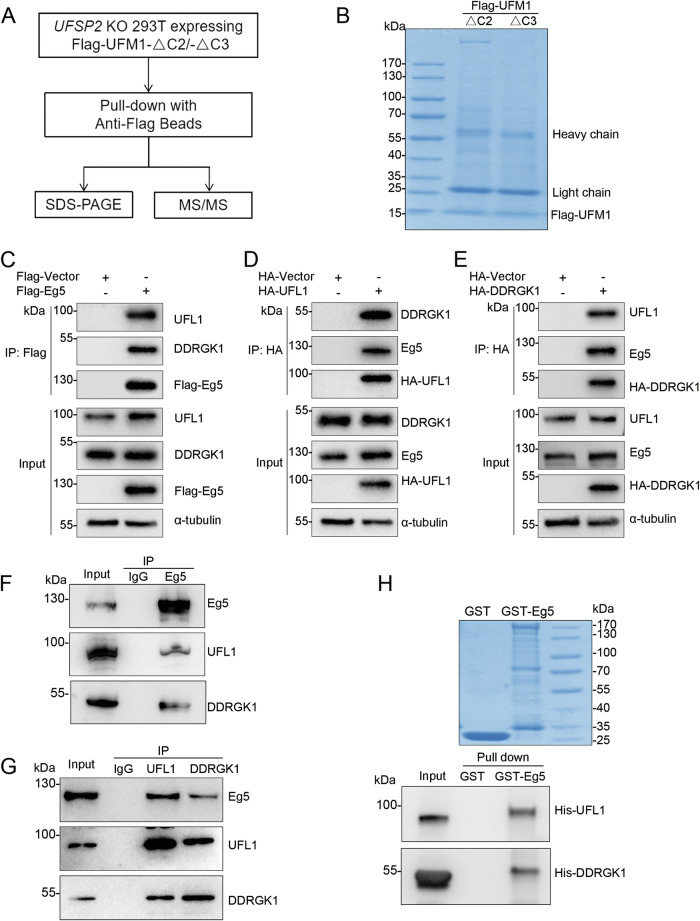
UFMylation is a highly conserved ubiquitin-like post-translational modification that catalyzes the covalent linkage of UFM1 to its target proteins. This modification plays a critical role in the maintenance of endoplasmic reticulum proteostasis, DNA damage response, autophagy, and transcriptional regulation. Mutations in UFM1, as well as in its specific E1 enzyme UBA5 and E2 enzyme UFC1, have been genetically linked to microcephaly. Our previous research unveiled the important role of UFMylation in regulating mitosis. However, the underlying mechanisms have remained unclear due to the limited identification of substrates. In this study, we identified Eg5, a motor protein crucial for mitotic spindle assembly and maintenance, as a novel substrate for UFMylation and identified Lys564 as the crucial UFMylation site. UFMylation did not alter its transcriptional level, phosphorylation level, or protein stability, but affected the mono-ubiquitination of Eg5. During mitosis, Eg5 and UFM1 co-localize at the centrosome and spindle apparatus, and defective UFMylation leads to diminished spindle localization of Eg5. Notably, the UFMylation-defective Eg5 mutant (K564R) exhibited shorter spindles, metaphase arrest, spindle checkpoint activation, and a failure of cell division in HeLa cells. Overall, Eg5 UFMylation is essential for proper spindle organization, mitotic progression, and cell proliferation.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






References
MeSH terms
Substances
Grants and funding
- 82071273/National Natural Science Foundation of China (National Science Foundation of China)
- 82271906/National Natural Science Foundation of China (National Science Foundation of China)
- 82101958/National Natural Science Foundation of China (National Science Foundation of China)
- 2019SK1010/Hunan Provincial Science and Technology Department (Department of Science and Technology of Hunan Province)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
