

Excitatory to inhibitory synaptic ratios are unchanged at presymptomatic stages in multiple models of ALS
- PMID: 39088455
- PMCID: PMC11293752
- DOI: 10.1371/journal.pone.0306423
Excitatory to inhibitory synaptic ratios are unchanged at presymptomatic stages in multiple models of ALS
Abstract
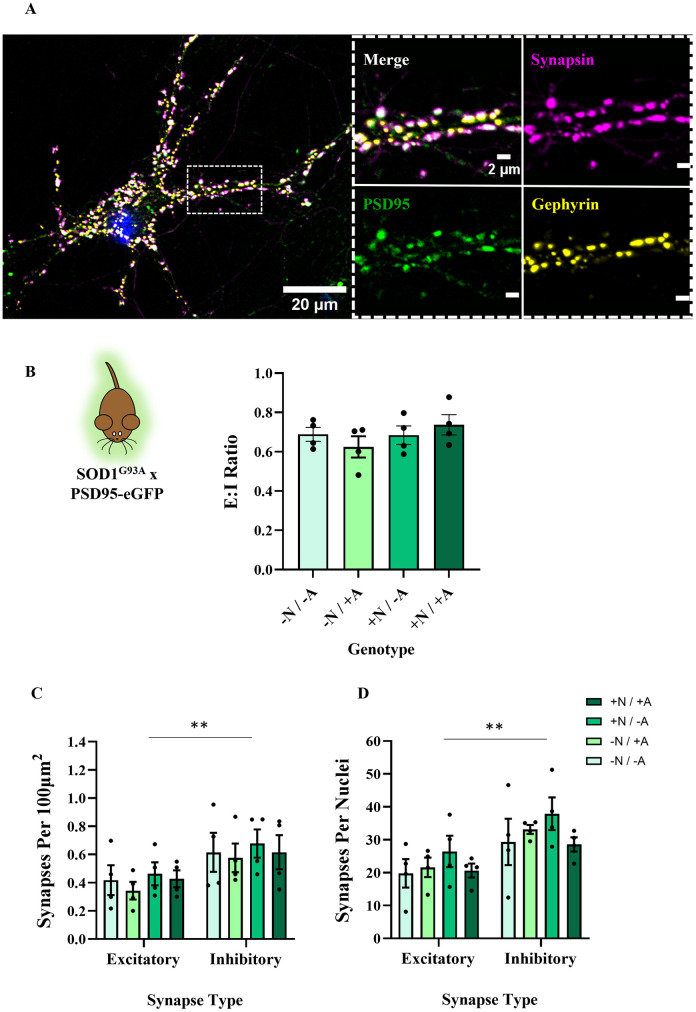
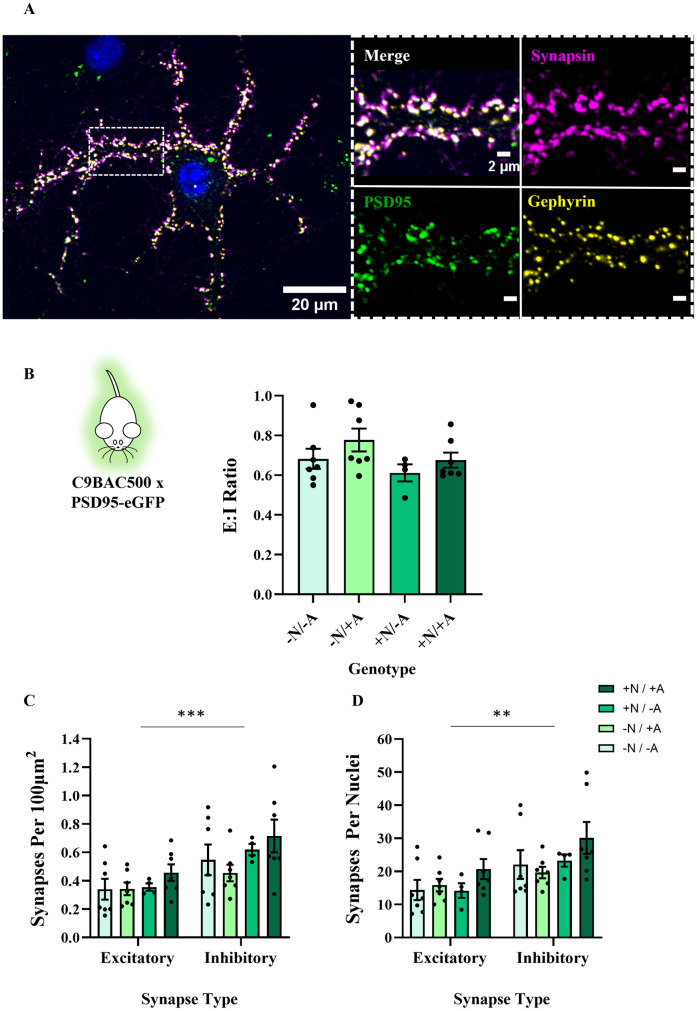
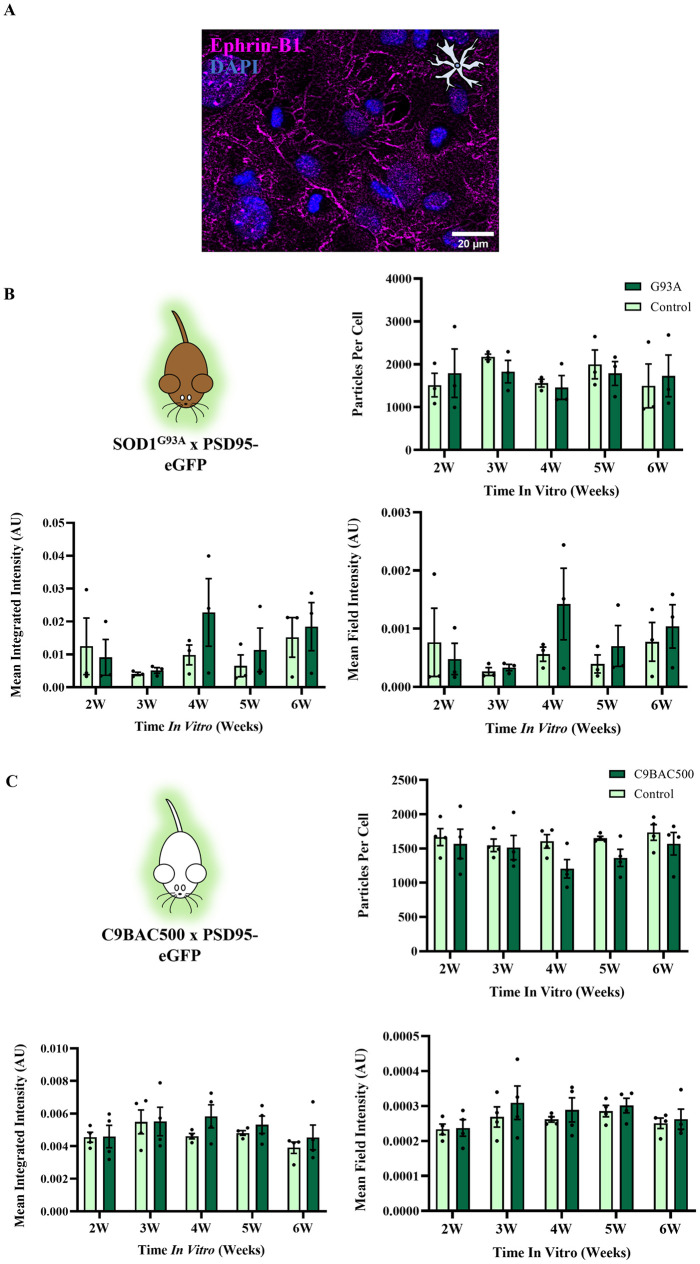
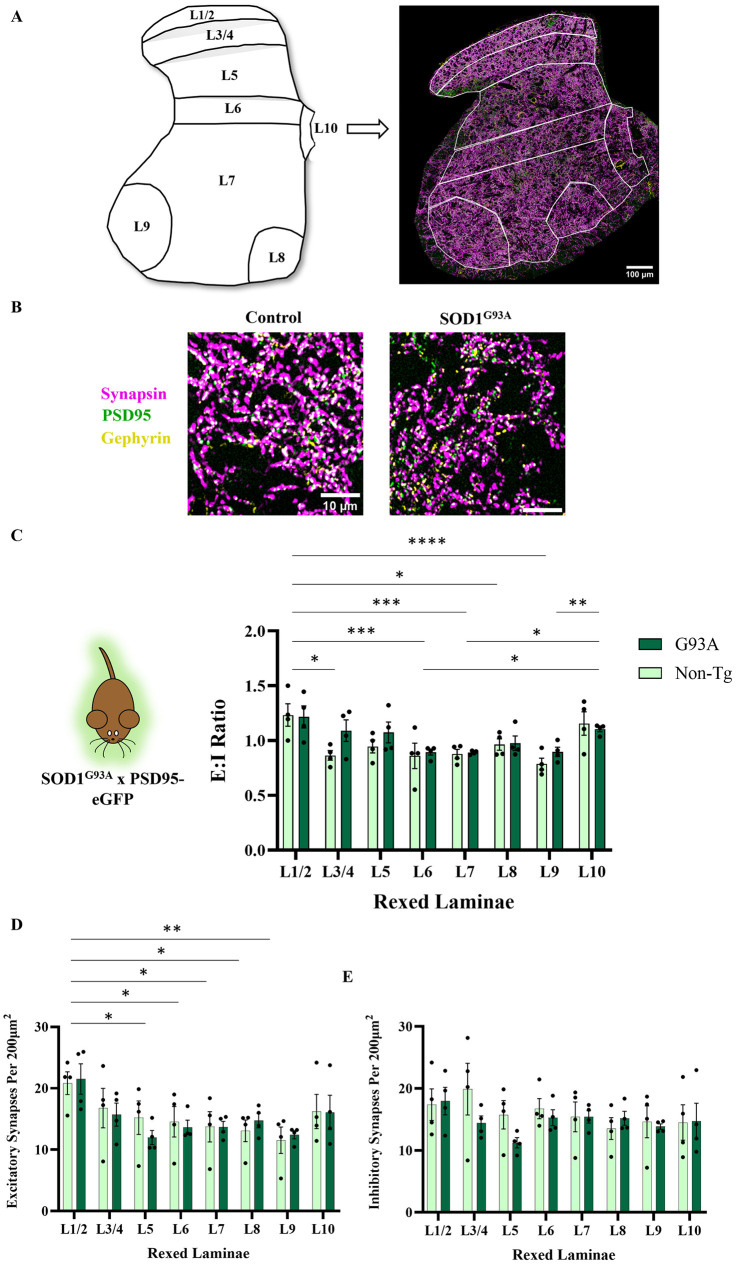
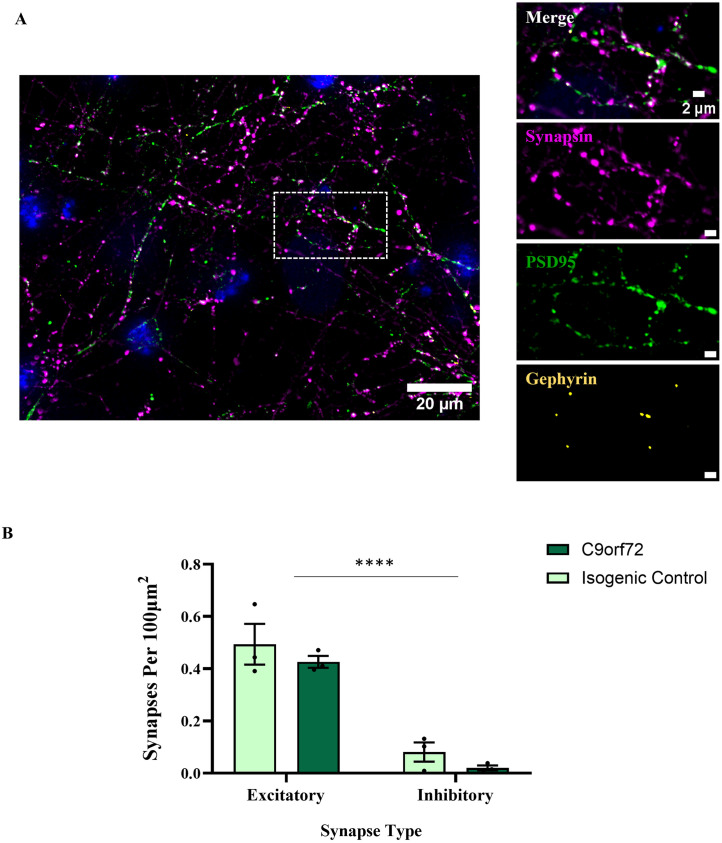
Hyperexcitability of motor neurons and spinal cord motor circuitry has been widely reported in the early stages of Amyotrophic Lateral Sclerosis (ALS). Changes in the relative amount of excitatory to inhibitory inputs onto a neuron (E:I synaptic ratio), possibly through a developmental shift in synapse formation in favour of excitatory transmission, could underlie pathological hyperexcitability. Given that astrocytes play a major role in early synaptogenesis and are implicated in ALS pathogenesis, their potential contribution to disease mechanisms involving synaptic imbalances and subsequent hyperexcitability is also of great interest. In order to assess E:I ratios in ALS, we utilised a novel primary spinal neuron / astrocyte co-culture system, derived from neonatal mice, in which synapses are formed in vitro. Using multiple ALS mouse models we found that no combination of astrocyte or neuron genotype produced alterations in E:I synaptic ratios assessed using pre- and post-synaptic anatomical markers. Similarly, we observed that ephrin-B1, a major contact-dependent astrocytic synaptogenic protein, was not differentially expressed by ALS primary astrocytes. Further to this, analysis of E:I ratios across the entire grey matter of the lumbar spinal cord in young (post-natal day 16-19) ALS mice revealed no differences versus controls. Finally, analysis in co-cultures of human iPSC-derived motor neurons and astrocytes harbouring the pathogenic C9orf72 hexanucleotide repeat expansion showed no evidence of a bias toward excitatory versus inhibitory synapse formation. We therefore conclude, utilising multiple ALS models, that we do not observe significant changes in the relative abundance of excitatory versus inhibitory synapses as would be expected if imbalances in synaptic inputs contribute to early hyperexcitability.
Copyright: © 2024 Bonthron et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- van Zundert B, Peuscher MH, Hynynen M, Chen A, Neve RL, Brown RH, et al.. Neonatal Neuronal Circuitry Shows Hyperexcitable Disturbance in a Mouse Model of the Adult-Onset Neurodegenerative Disease Amyotrophic Lateral Sclerosis. Journal of Neuroscience. 2008;28: 10864–10874. doi: 10.1523/JNEUROSCI.1340-08.2008 - DOI - PMC - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
