

This is a preprint.
Prdm16-dependent antigen-presenting cells induce tolerance to intestinal antigens
- PMID: 39091750
- PMCID: PMC11291166
- DOI: 10.1101/2024.07.23.604803
Prdm16-dependent antigen-presenting cells induce tolerance to intestinal antigens
Update in
-
PRDM16-dependent antigen-presenting cells induce tolerance to gut antigens.Nature. 2025 Jun;642(8068):756-765. doi: 10.1038/s41586-025-08982-4. Epub 2025 Apr 14. Nature. 2025. PMID: 40228524 Free PMC article.
Abstract
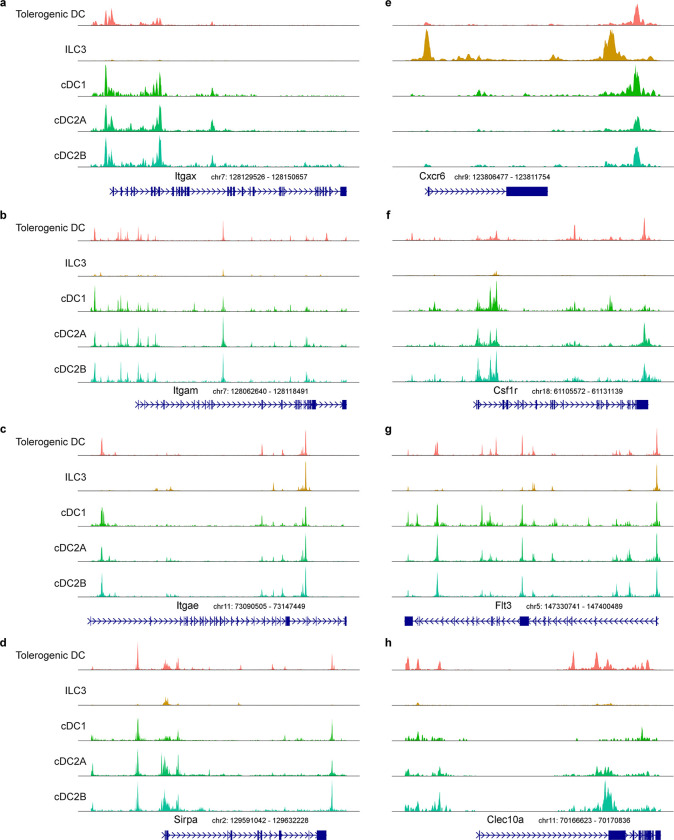
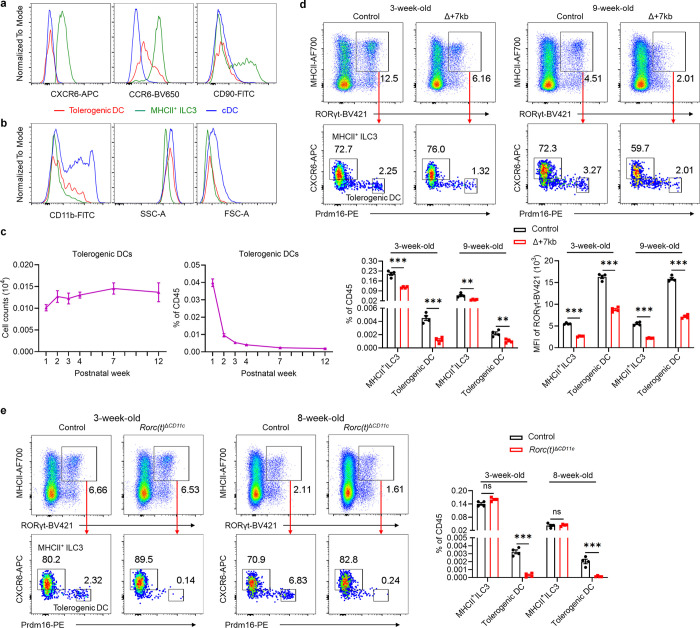
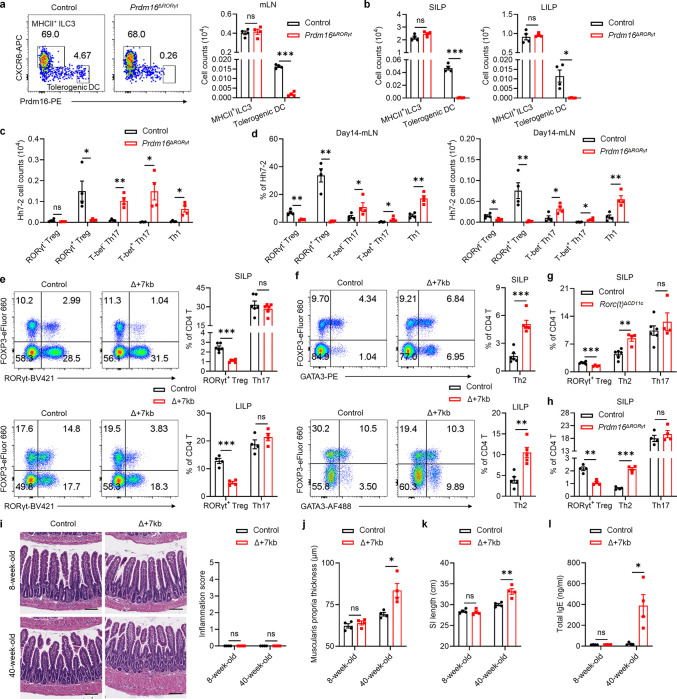
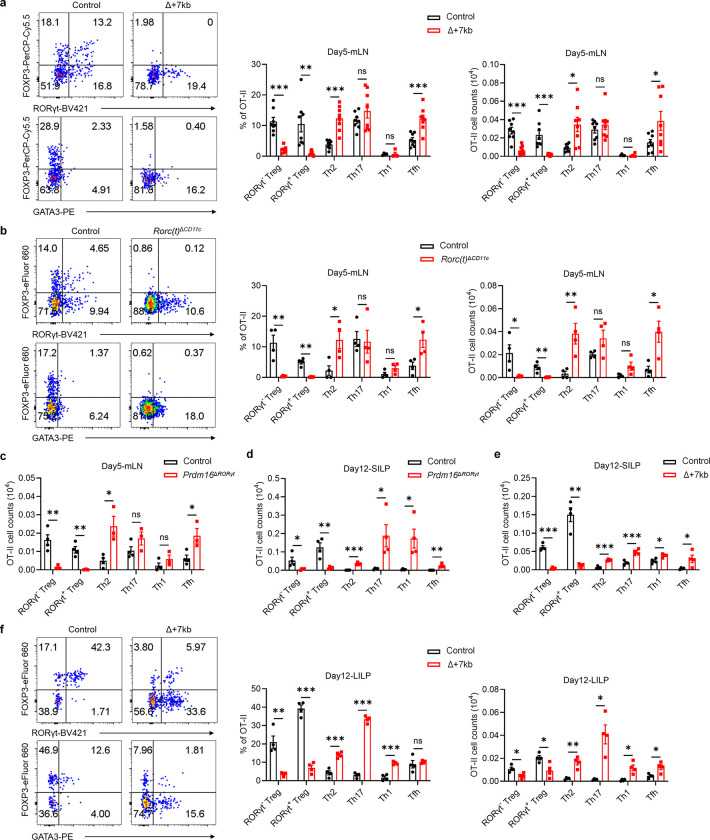
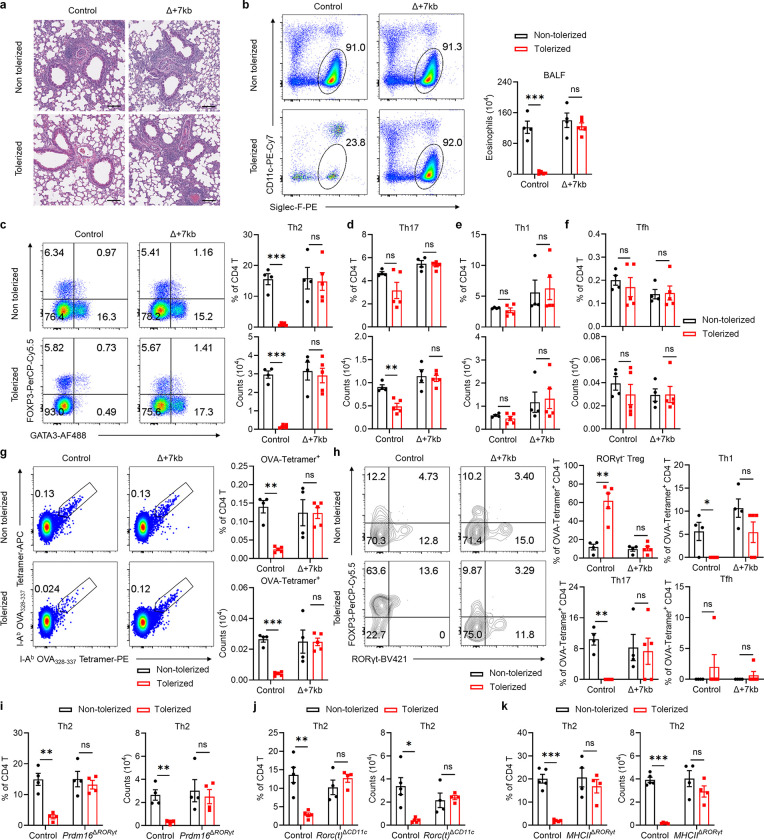
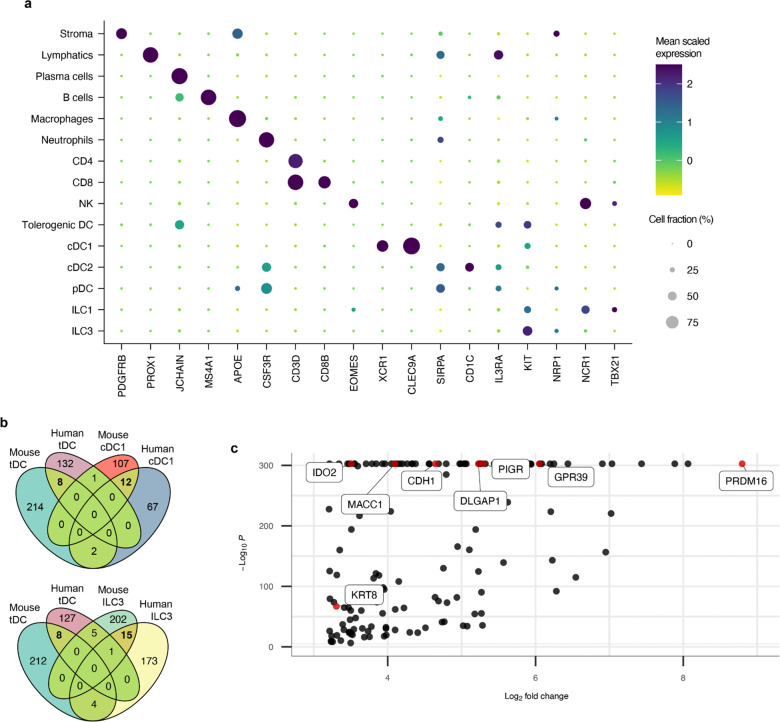
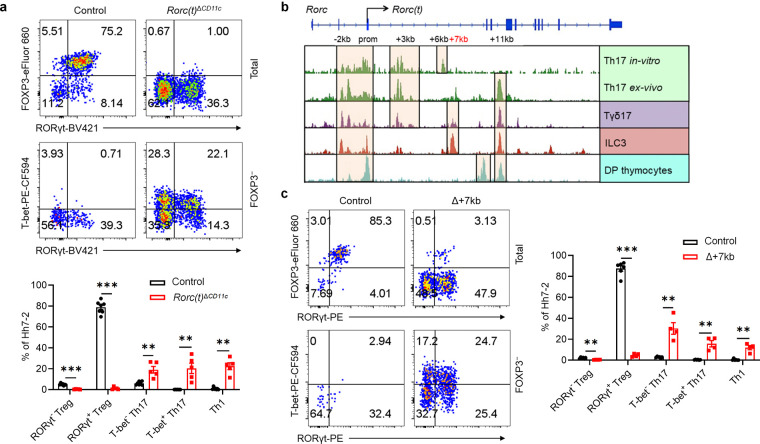
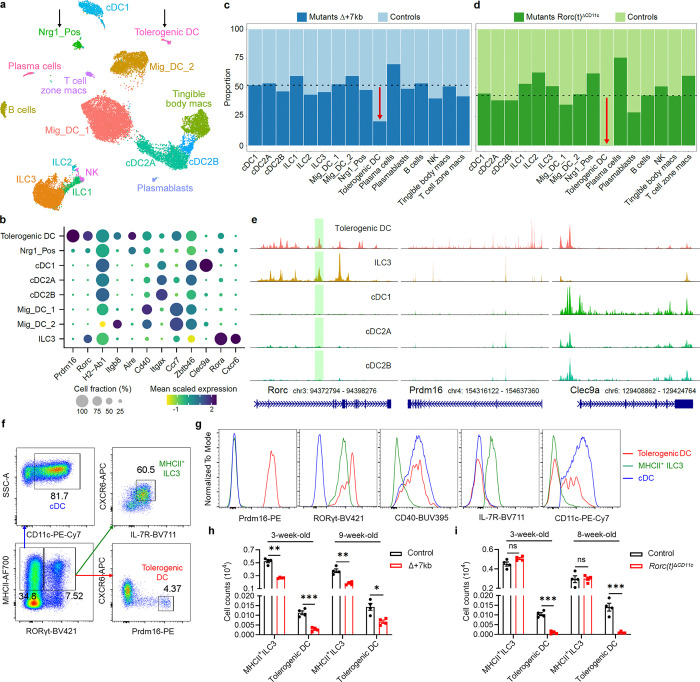
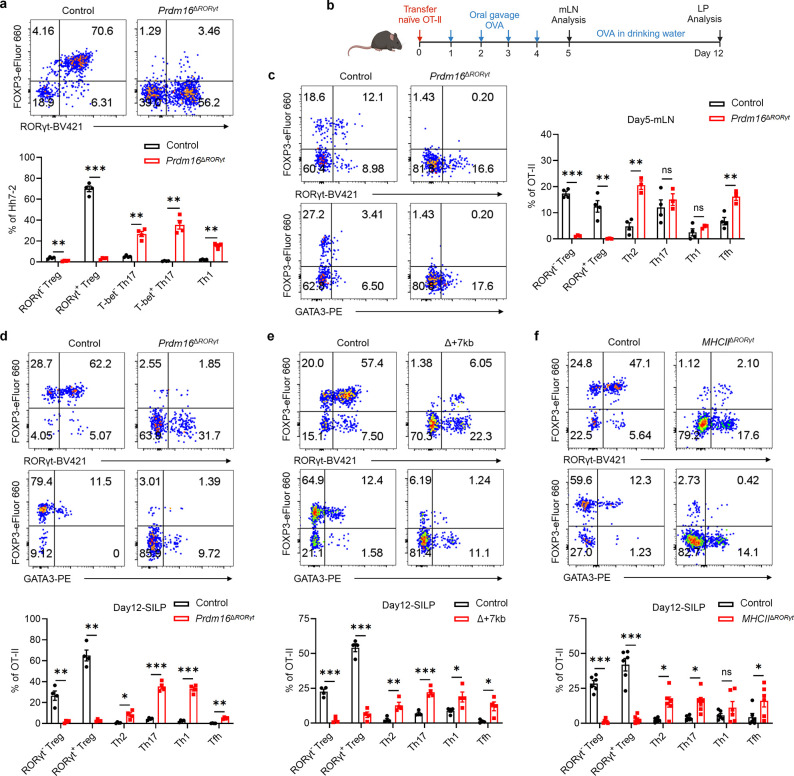
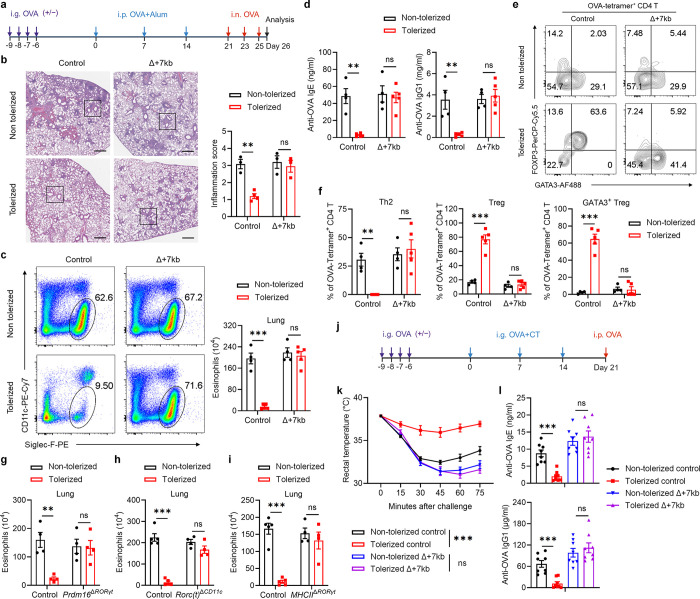
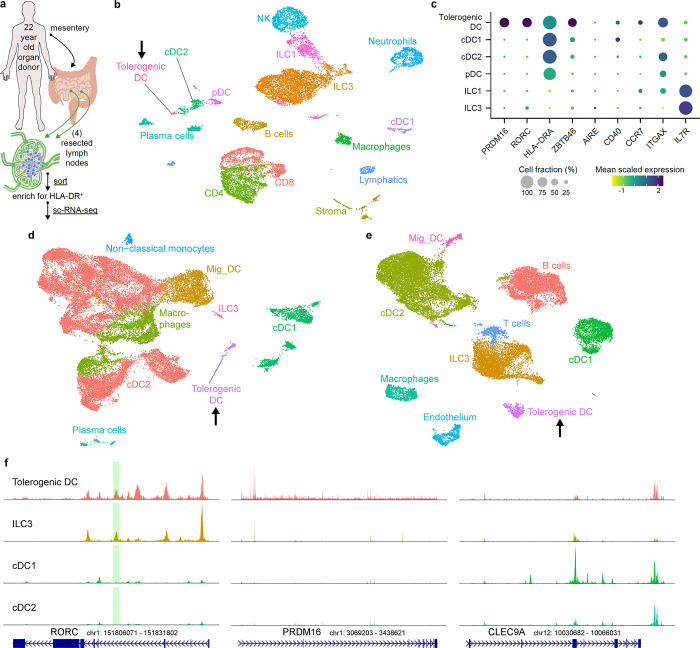
The gastrointestinal tract is continuously exposed to foreign antigens in food and commensal microbes with potential to induce adaptive immune responses. Peripherally induced T regulatory (pTreg) cells are essential for mitigating inflammatory responses to these agents1-4. While RORγt+ antigen-presenting cells (RORγt-APCs) were shown to program gut microbiota-specific pTreg5-7, their definition remains incomplete, and the APC responsible for food tolerance has remained elusive. Here, we identify a distinct subset of RORγt-APCs, designated tolerogenic dendritic cells (tDC), required for differentiation of both food- and microbiota-specific pTreg cells and for establishment of oral tolerance. tDC development and function require expression of the transcription factors Prdm16 and RORγt, as well as a unique Rorc(t) cis-regulatory element. Gene expression, chromatin accessibility, and surface marker analysis establish tDC as myeloid in origin, distinct from ILC3, and sharing epigenetic profiles with classical DC. Upon genetic perturbation of tDC, we observe a substantial increase in food antigen-specific T helper 2 (Th2) cells in lieu of pTreg, leading to compromised tolerance in mouse models of asthma and food allergy. Single-cell analyses of freshly resected mesenteric lymph nodes from a human organ donor, as well as multiple specimens of human intestine and tonsil, reveal candidate tDC with co-expression of PRDM16 and RORC and an extensive transcriptome shared with mice, highlighting an evolutionarily conserved role across species. Our findings suggest that a better understanding of how tDC develop and how they regulate T cell responses to food and microbial antigens could offer new insights into developing therapeutic strategies for autoimmune and allergic diseases as well as organ transplant tolerance.
Conflict of interest statement
Competing interests D.R.L. is cofounder of Vedanta Biosciences and ImmunAI, on the advisory boards of IMIDomics, Sonoma Biotherapeutics, NILO Therapeutics, and Evommune, and on the board of directors of Pfizer Inc. All other authors declare no competing interests.
Figures
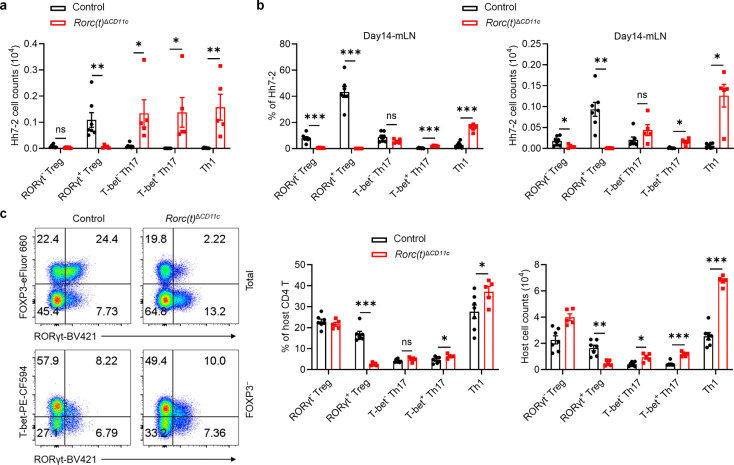
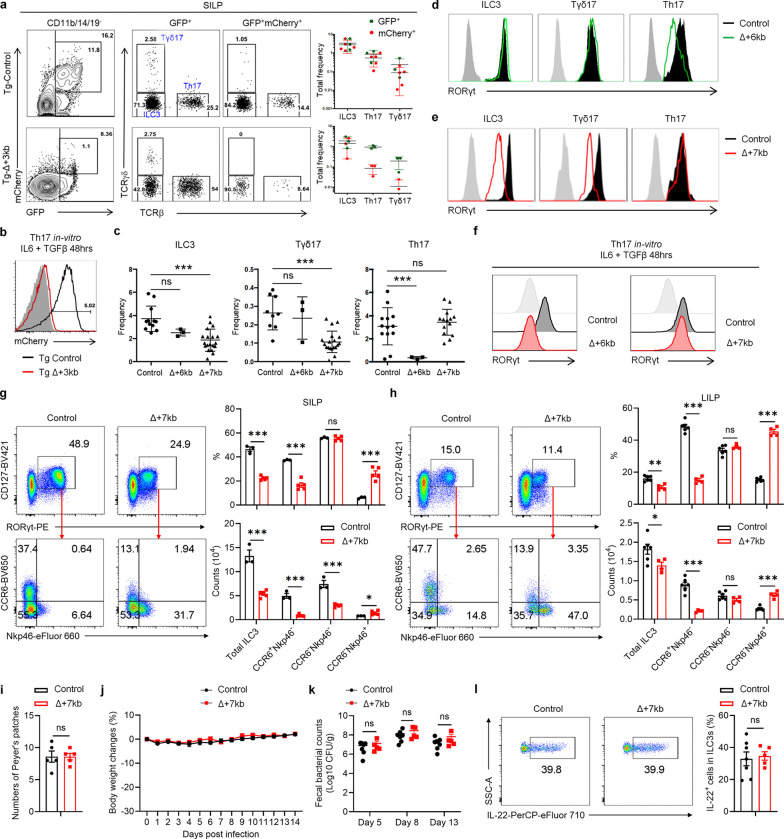
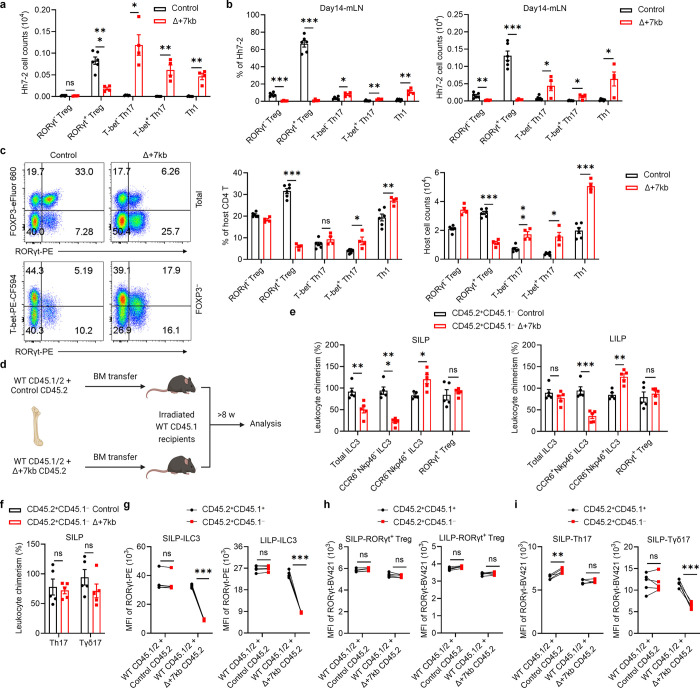
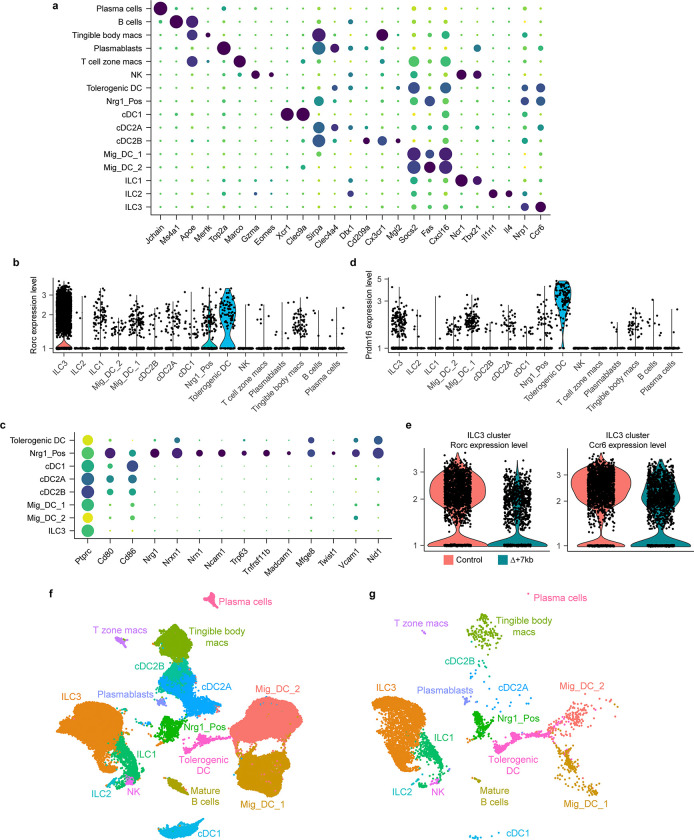
References
-
- Bilate A. M. & Lafaille J. J. Induced CD4+Foxp3+ regulatory T cells in immune tolerance. Annu. Rev. Immunol. 30, 733–758 (2012). - PubMed
-
- Whibley N., Tucci A. & Powrie F. Regulatory T cell adaptation in the intestine and skin. Nat. Immunol. 20, 386–396 (2019). - PubMed
-
- Ramanan D. et al. Regulatory T cells in the face of the intestinal microbiota. Nat. Rev. Immunol. 23, 749–762 (2023). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous