

This is a preprint.
An ultraconserved snoRNA-like element in long noncoding RNA CRNDE promotes ribosome biogenesis and cell proliferation
- PMID: 39091767
- PMCID: PMC11291168
- DOI: 10.1101/2024.07.23.604857
An ultraconserved snoRNA-like element in long noncoding RNA CRNDE promotes ribosome biogenesis and cell proliferation
Update in
-
An ultraconserved snoRNA-like element in long noncoding RNA CRNDE promotes ribosome biogenesis and cell proliferation.Mol Cell. 2025 Apr 17;85(8):1543-1560.e10. doi: 10.1016/j.molcel.2025.03.006. Epub 2025 Apr 3. Mol Cell. 2025. PMID: 40185099
Abstract
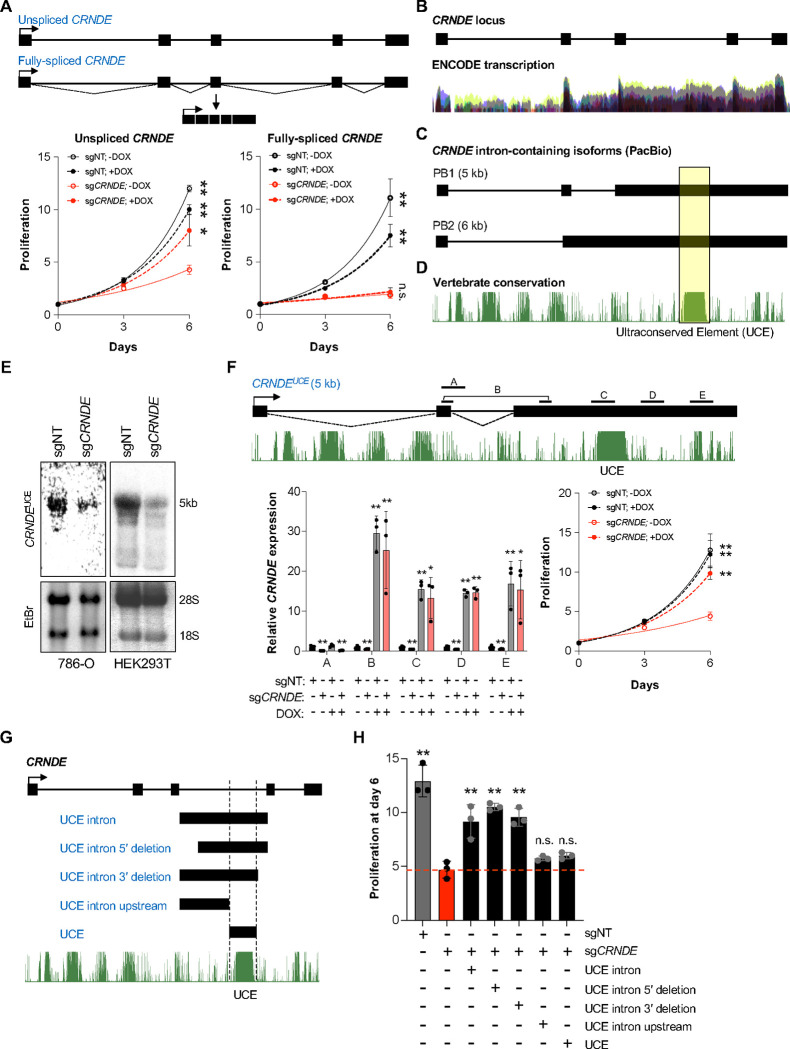
Cancer cells frequently upregulate ribosome production to support tumorigenesis. While small nucleolar RNAs (snoRNAs) are critical for ribosome biogenesis, the roles of other classes of noncoding RNAs in this process remain largely unknown. Here we performed CRISPRi screens to identify essential long noncoding RNAs (lncRNAs) in renal cell carcinoma (RCC) cells. This revealed that an alternatively-spliced isoform of lncRNA Colorectal Neoplasia Differentially Expressed containing an ultraconserved element (UCE), referred to as CRNDE UCE, is required for RCC cell proliferation. CRNDE UCE localizes to the nucleolus and promotes 60S ribosomal subunit biogenesis. The UCE of CRNDE functions as an unprocessed C/D box snoRNA that directly interacts with ribosomal RNA precursors. This facilitates delivery of eIF6, a key 60S biogenesis factor, which binds to CRNDE UCE through a sequence element adjacent to the UCE. These findings highlight the functional versatility of snoRNA sequences and expand the known mechanisms through which noncoding RNAs orchestrate ribosome biogenesis.
Conflict of interest statement
DECLARATION OF INTERESTS J.T.M is a scientific advisor for Ribometrix, Inc. and owns equity in Orbital Therapeutics, Inc.
Figures





References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources