

This is a preprint.
Meiosis-specific decoupling of the pericentromere from the kinetochore
- PMID: 39091844
- PMCID: PMC11291024
- DOI: 10.1101/2024.07.21.604490
Meiosis-specific decoupling of the pericentromere from the kinetochore
Abstract
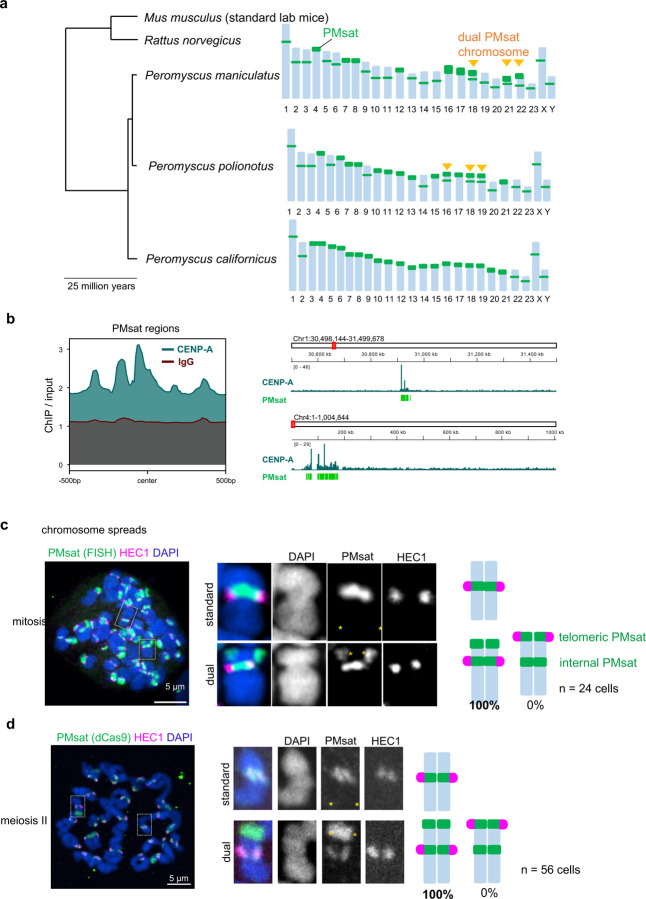
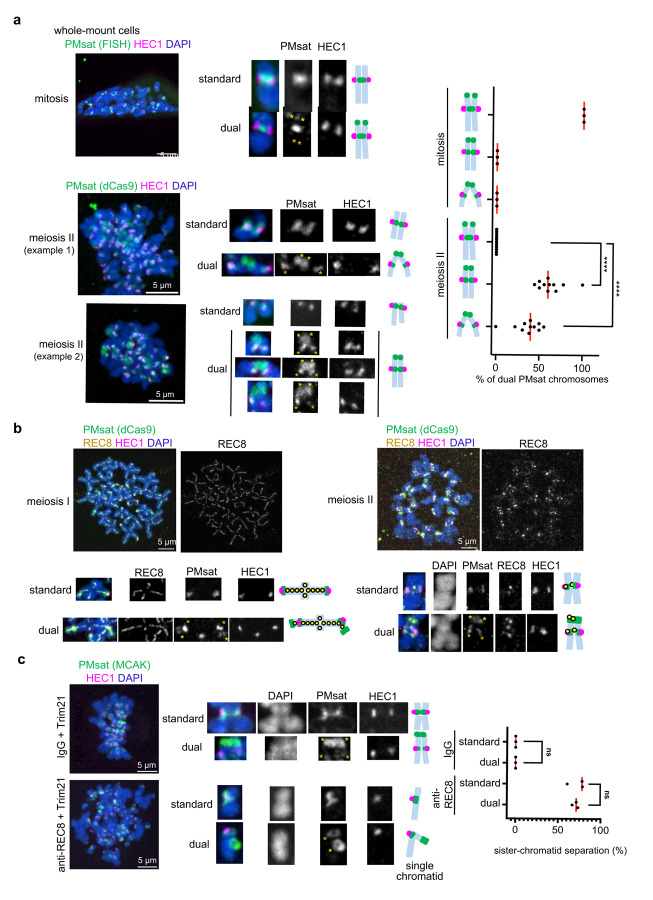
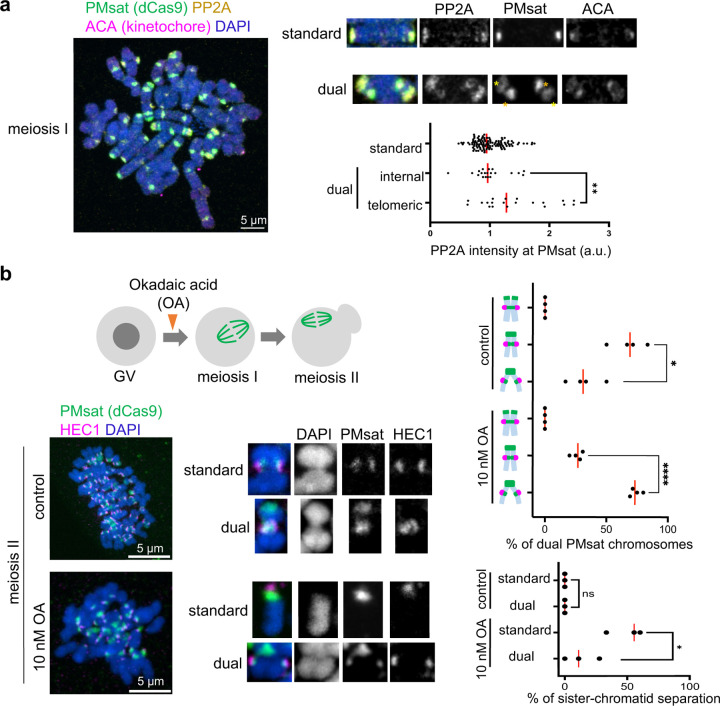
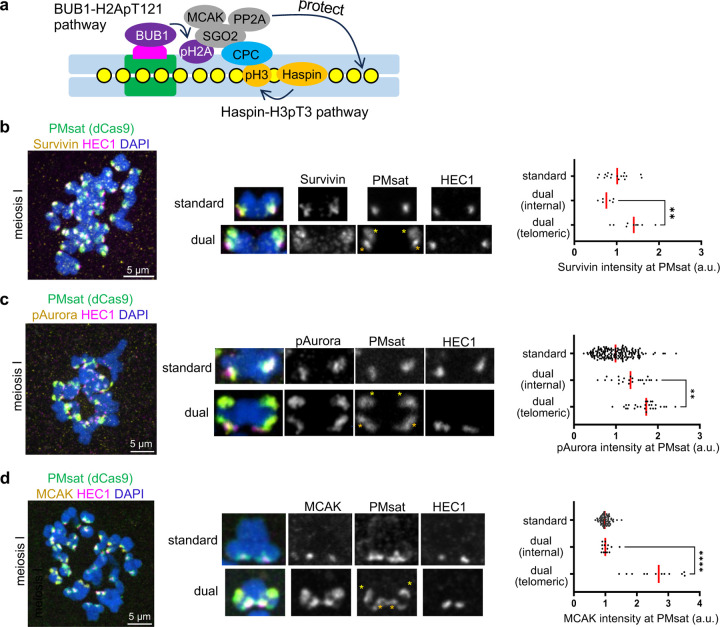
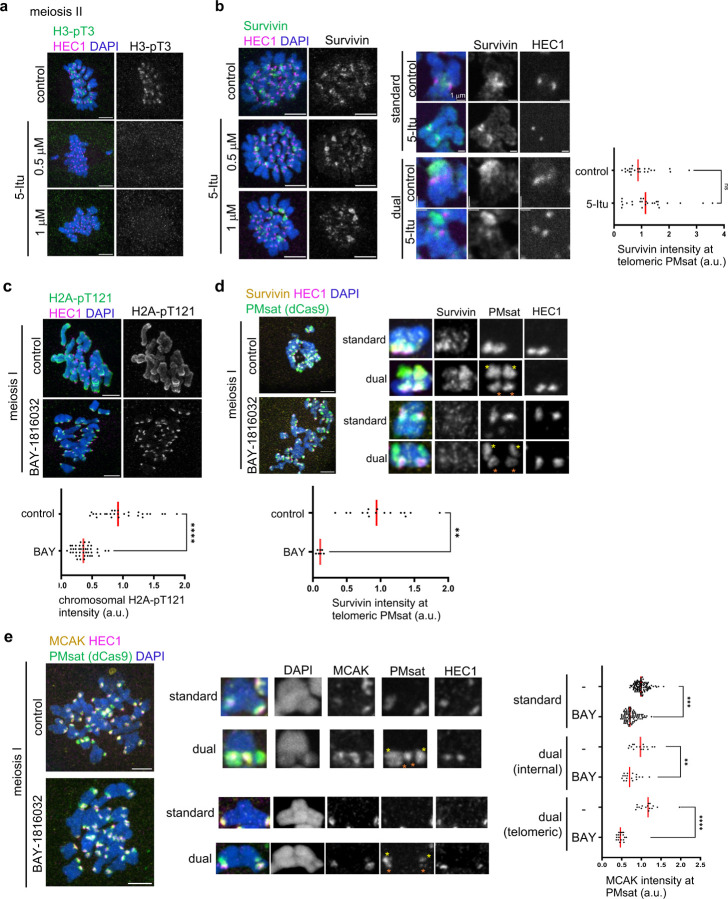
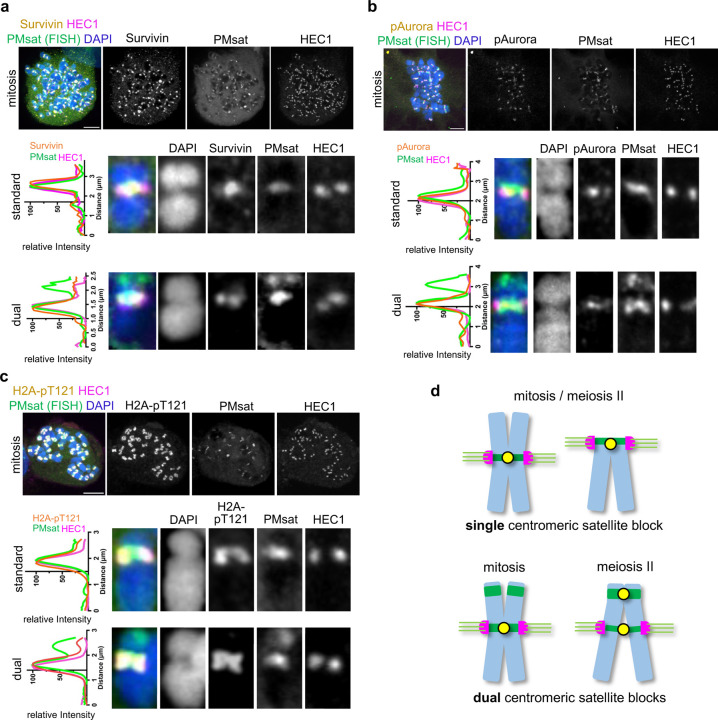
The primary constriction site of the M-phase chromosome is an established marker for the kinetochore position, often used to determine the karyotype of each species. Underlying this observation is the concept that the kinetochore is spatially linked with the pericentromere where sister-chromatids are most tightly cohered. Here, we found an unconventional pericentromere specification with sister chromatids mainly cohered at a chromosome end, spatially separated from the kinetochore in Peromyscus mouse oocytes. This distal locus enriched cohesin protectors, such as the Chromosomal Passenger Complex (CPC) and PP2A, at a higher level compared to its centromere/kinetochore region, acting as the primary site for sister-chromatid cohesion. Chromosomes with the distal cohesion site exhibited enhanced cohesin protection at anaphase I compared to those without it, implying that these distal cohesion sites may have evolved to ensure sister-chromatid cohesion during meiosis. In contrast, mitotic cells enriched CPC only near the kinetochore and the distal locus was not cohered between sister chromatids, suggesting a meiosis-specific mechanism to protect cohesin at this distal locus. We found that this distal locus corresponds to an additional centromeric satellite block, located far apart from the centromeric satellite block that builds the kinetochore. Several Peromyscus species carry chromosomes with two such centromeric satellite blocks. Analyses on three Peromyscus species revealed that the internal satellite consistently assembles the kinetochore in both mitosis and meiosis, whereas the distal satellite selectively enriches cohesin protectors in meiosis to promote sister-chromatid cohesion at that site. Thus, our study demonstrates that pericentromere specification is remarkably flexible and can control chromosome segregation in a cell-type and context dependent manner.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Cheeseman I. M. & Desai A. Molecular architecture of the kinetochore-microtubule interface. Nature reviews. Molecular cell biology 9, 33–46 (2008). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources