

Using Organoids to Model Sex Differences in the Human Brain
- PMID: 39092139
- PMCID: PMC11292257
- DOI: 10.1016/j.bpsgos.2024.100343
Using Organoids to Model Sex Differences in the Human Brain
Abstract
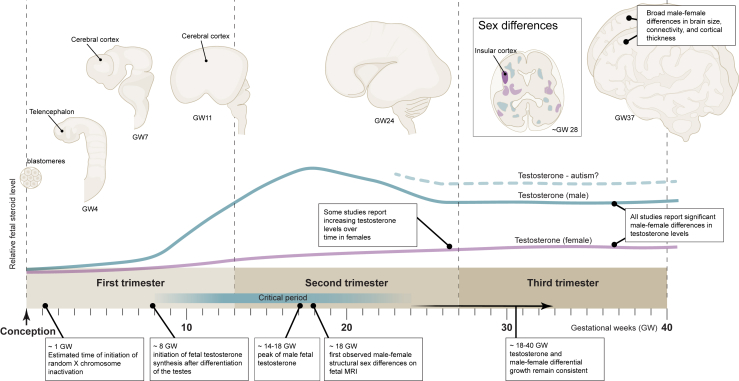
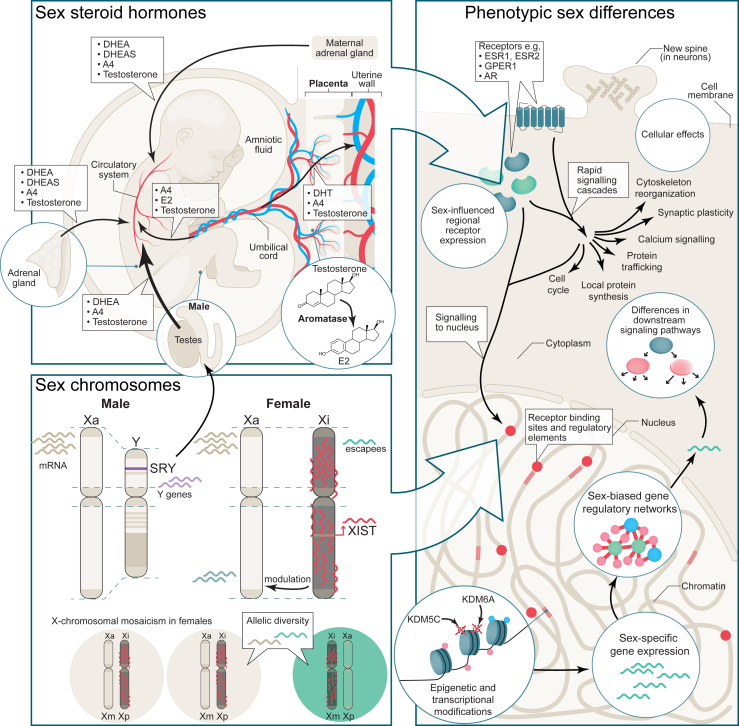
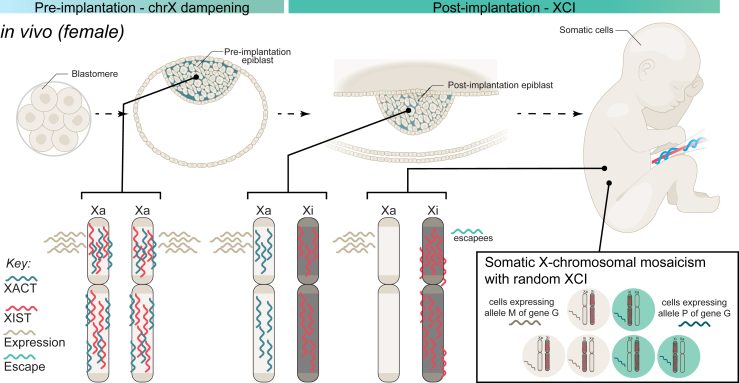
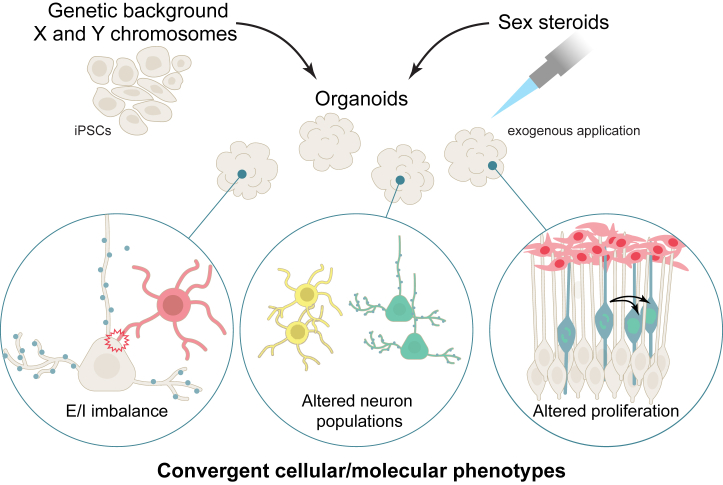
Sex differences are widespread during neurodevelopment and play a role in neuropsychiatric conditions such as autism, which is more prevalent in males than females. In humans, males have been shown to have larger brain volumes than females with development of the hippocampus and amygdala showing prominent sex differences. Mechanistically, sex steroids and sex chromosomes drive these differences in brain development, which seem to peak during prenatal and pubertal stages. Animal models have played a crucial role in understanding sex differences, but the study of human sex differences requires an experimental model that can recapitulate complex genetic traits. To fill this gap, human induced pluripotent stem cell-derived brain organoids are now being used to study how complex genetic traits influence prenatal brain development. For example, brain organoids from individuals with autism and individuals with X chromosome-linked Rett syndrome and fragile X syndrome have revealed prenatal differences in cell proliferation, a measure of brain volume differences, and excitatory-inhibitory imbalances. Brain organoids have also revealed increased neurogenesis of excitatory neurons due to androgens. However, despite growing interest in using brain organoids, several key challenges remain that affect its validity as a model system. In this review, we discuss how sex steroids and the sex chromosomes each contribute to sex differences in brain development. Then, we examine the role of X chromosome inactivation as a factor that drives sex differences. Finally, we discuss the combined challenges of modeling X chromosome inactivation and limitations of brain organoids that need to be taken into consideration when studying sex differences.
Keywords: Autism; Brain organoids; Sex chromosomes; Sex differences; Steroids; X chromosome inactivation.
Plain language summary
Sex differences are a contributing factor in neuropsychiatric conditions such as autism, which is more prevalent in males. Sex differences occur through interactions between sex steroid hormones such as estrogen and testosterone and sex chromosomes (chrX and chrY). Human stem cell–derived brain organoids are laboratory models that mimic brain development. For example, in individuals with neurodevelopmental conditions, brain organoids have revealed an imbalance of neuron populations compared with neurotypical individuals. In this review, we discuss sex steroid and sex chromosome influences on brain development and challenges of this model that need to be taken into account when studying sex differences.
© 2024 Published by Elsevier Inc on behalf of Society of Biological Psychiatry.
Figures
References
-
- Dewing P., Shi T., Horvath S., Vilain E. Sexually dimorphic gene expression in mouse brain precedes gonadal differentiation. Brain Res Mol Brain Res. 2003;118:82–90. - PubMed
-
- Wierman M.E. Sex steroid effects at target tissues: Mechanisms of action. Adv Physiol Educ. 2007;31:26–33. - PubMed
-
- Alexander G.M., Wilcox T., Woods R. Sex differences in infants’ visual interest in toys. Arch Sex Behav. 2009;38:427–433. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous
