

Activation of the helper NRC4 immune receptor forms a hexameric resistosome
- PMID: 39094568
- PMCID: PMC11380581
- DOI: 10.1016/j.cell.2024.07.013
Activation of the helper NRC4 immune receptor forms a hexameric resistosome
Abstract
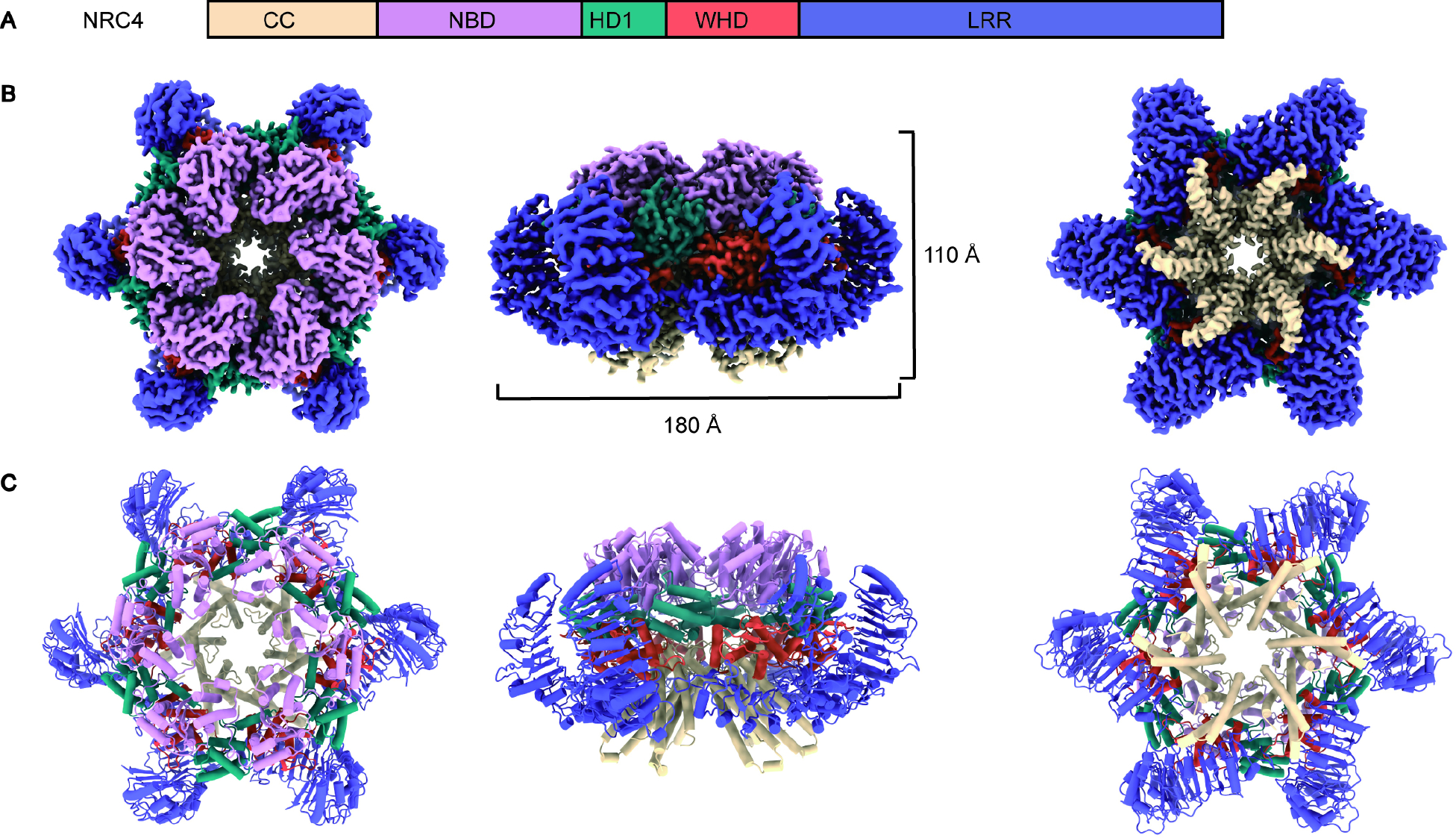
Innate immune responses to microbial pathogens are regulated by intracellular receptors known as nucleotide-binding leucine-rich repeat receptors (NLRs) in both the plant and animal kingdoms. Across plant innate immune systems, "helper" NLRs (hNLRs) work in coordination with "sensor" NLRs (sNLRs) to modulate disease resistance signaling pathways. Activation mechanisms of hNLRs based on structures are unknown. Our research reveals that the hNLR, known as NLR required for cell death 4 (NRC4), assembles into a hexameric resistosome upon activation by the sNLR Bs2 and the pathogenic effector AvrBs2. This conformational change triggers immune responses by facilitating the influx of calcium ions (Ca2+) into the cytosol. The activation mimic alleles of NRC2, NRC3, or NRC4 alone did not induce Ca2+ influx and cell death in animal cells, suggesting that unknown plant-specific factors regulate NRCs' activation in plants. These findings significantly advance our understanding of the regulatory mechanisms governing plant immune responses.
Keywords: ETI; NLR proteins; NRC0; NRC2; NRC3; NRC4 resistosome; PTI; calcium influx; pathogen recognition; plant immunity.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests B.J.S. is a scientific co-founder of and serves on the board of directors for Mendel Biotechnology. He also serves on the scientific advisory boards of Verinomics and the Sainsbury Laboratory.
Figures





Update of
-
The activated plant NRC4 immune receptor forms a hexameric resistosome.bioRxiv [Preprint]. 2024 Jan 9:2023.12.18.571367. doi: 10.1101/2023.12.18.571367. bioRxiv. 2024. Update in: Cell. 2024 Sep 5;187(18):4877-4889.e15. doi: 10.1016/j.cell.2024.07.013. PMID: 38187616 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
