

Mass vaccination with reassortment-impaired live H9N2 avian influenza vaccine
- PMID: 39097573
- PMCID: PMC11297921
- DOI: 10.1038/s41541-024-00923-y
Mass vaccination with reassortment-impaired live H9N2 avian influenza vaccine
Abstract
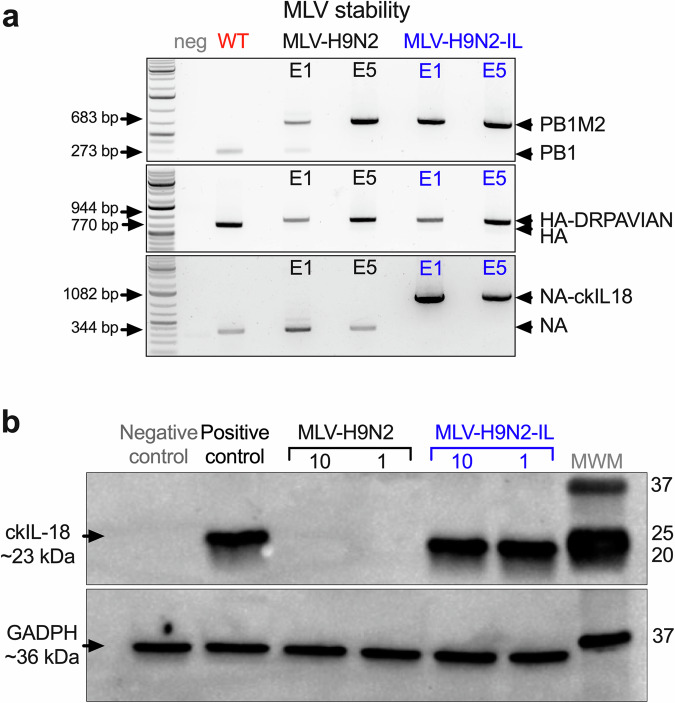
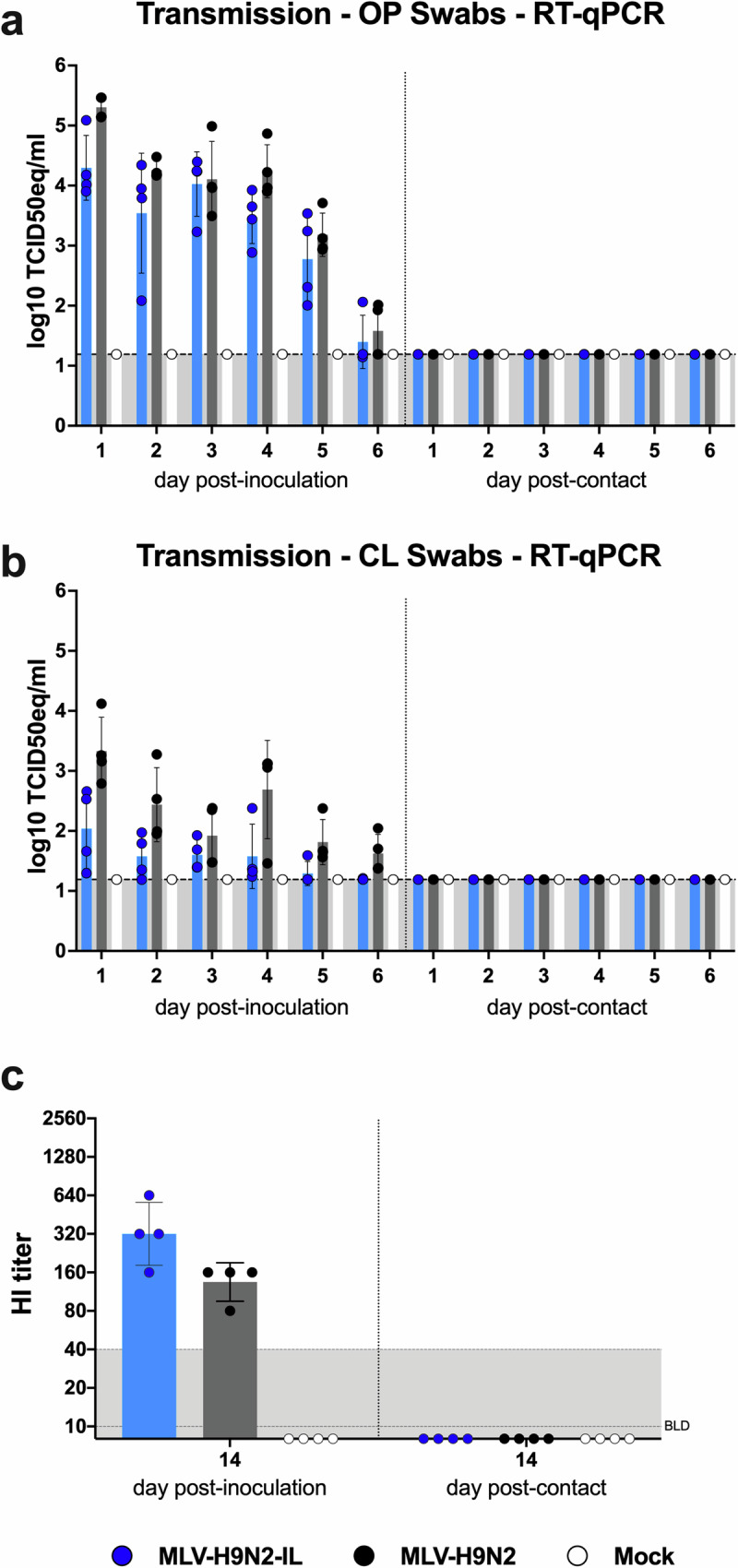
Avian influenza poses a severe threat to poultry production and global food security, prompting the development of vaccination programs in numerous countries. Modified live virus (MLV) vaccines, with their potential for mass application, offer a distinct advantage over existing options. However, concerns surrounding reversion, recombination, and unintended transmission have hindered the progress of MLV development for avian influenza in poultry. To address these concerns, we engineered reassortment-impaired, non-transmissible, safe, immunogenic, and protective MLVs through the rearrangement of internal gene segments and additional modifications to the surface gene segments HA and NA. The unique peptide marker aspartic acid-arginine-proline-alanine-valine-isoleucine-alanine-asparragine (DRPAVIAN) was incorporated into HA, while NA was modified to encode the chicken interleukin-18 (ckIL18) gene (MLV-H9N2-IL). In vitro, the MLV-H9N2 and MLV-H9N2-IL candidates demonstrated stability and virus titers comparable to the wild-type H9N2 strain. In chickens, the MLV-H9N2 and MLV-H9N2-IL candidates did not transmit via direct contact. Co-infection studies with wild-type virus confirmed that the altered HA and NA segments exhibited fitness disadvantages and did not reassort. Vaccinated chickens showed no clinical signs upon vaccination, all seroconverted, and the inclusion of ckIL18 in the MLV-H9N2-IL vaccine enhanced neutralizing antibody production. A significant decrease in viral loads post-challenge underscored the protective effect of the MLVs. The MLV-H9N2-IL vaccine, administered via drinking water, proved immunogenic in chickens in a dose-dependent manner, generating protective levels of neutralizing antibodies upon aggressive homologous virus challenge. In summary, this study lays the groundwork for safe MLVs against avian influenza suitable for mass vaccination efforts.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Grants and funding
LinkOut - more resources
Full Text Sources
