

Metabolic adaptations underpin high productivity rates in relict subsurface water
- PMID: 39103408
- PMCID: PMC11300587
- DOI: 10.1038/s41598-024-68868-9
Metabolic adaptations underpin high productivity rates in relict subsurface water
Abstract
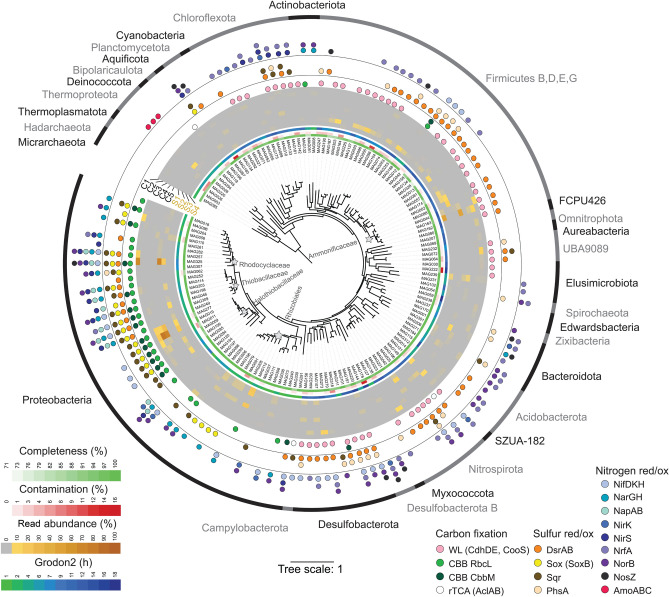
Groundwater aquifers are ecological hotspots with diverse microbes essential for biogeochemical cycles. Their ecophysiology has seldom been studied on a basin scale. In particular, our knowledge of chemosynthesis in the deep aquifers where temperatures reach 60 °C, is limited. Here, we investigated the diversity, activity, and metabolic potential of microbial communities from nine wells reaching ancient groundwater beneath Israel's Negev Desert, spanning two significant, deep (up to 1.5 km) aquifers, the Judea Group carbonate and Kurnub Group Nubian sandstone that contain fresh to brackish, hypoxic to anoxic water. We estimated chemosynthetic productivity rates ranging from 0.55 ± 0.06 to 0.82 ± 0.07 µg C L-1 d-1 (mean ± SD), suggesting that aquifer productivity may be underestimated. We showed that 60% of MAGs harbored genes for autotrophic pathways, mainly the Calvin-Benson-Bassham cycle and the Wood-Ljungdahl pathway, indicating a substantial chemosynthetic capacity within these microbial communities. We emphasize the potential metabolic versatility in the deep subsurface, enabling efficient carbon and energy use. This study set a precedent for global aquifer exploration, like the Nubian Sandstone Aquifer System in the Arabian and Western Deserts, and reconsiders their role as carbon sinks.
Keywords: Ancient groundwater; Carbon fixation; Deep terrestrial subsurface; Metagenomics; Primary production.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Magnabosco, C. et al. The biomass and biodiversity of the continental subsurface. Nat. Geosci.11(10), 707–717. 10.1038/s41561-018-0221-6 (2018). 10.1038/s41561-018-0221-6 - DOI
-
- Ragon, M., Van Driessche, A. E. S., García-Ruíz, J. M., Moreira, D. & López-García, P. Microbial diversity in the deep-subsurface hydrothermal aquifer feeding the giant gypsum crystal-bearing Naica Mine Mexico. Front. Microbiol.4, 37. 10.3389/fmicb.2013.00037 (2013). 10.3389/fmicb.2013.00037 - DOI - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
