

Reprogramming the tumor immune microenvironment using engineered dual-drug loaded Salmonella
- PMID: 39107284
- PMCID: PMC11303714
- DOI: 10.1038/s41467-024-50950-5
Reprogramming the tumor immune microenvironment using engineered dual-drug loaded Salmonella
Erratum in
-
Author Correction: Reprogramming the tumor immune microenvironment using engineered dual-drug loaded Salmonella.Nat Commun. 2025 Jan 27;16(1):1084. doi: 10.1038/s41467-025-56044-0. Nat Commun. 2025. PMID: 39870640 Free PMC article. No abstract available.
Abstract
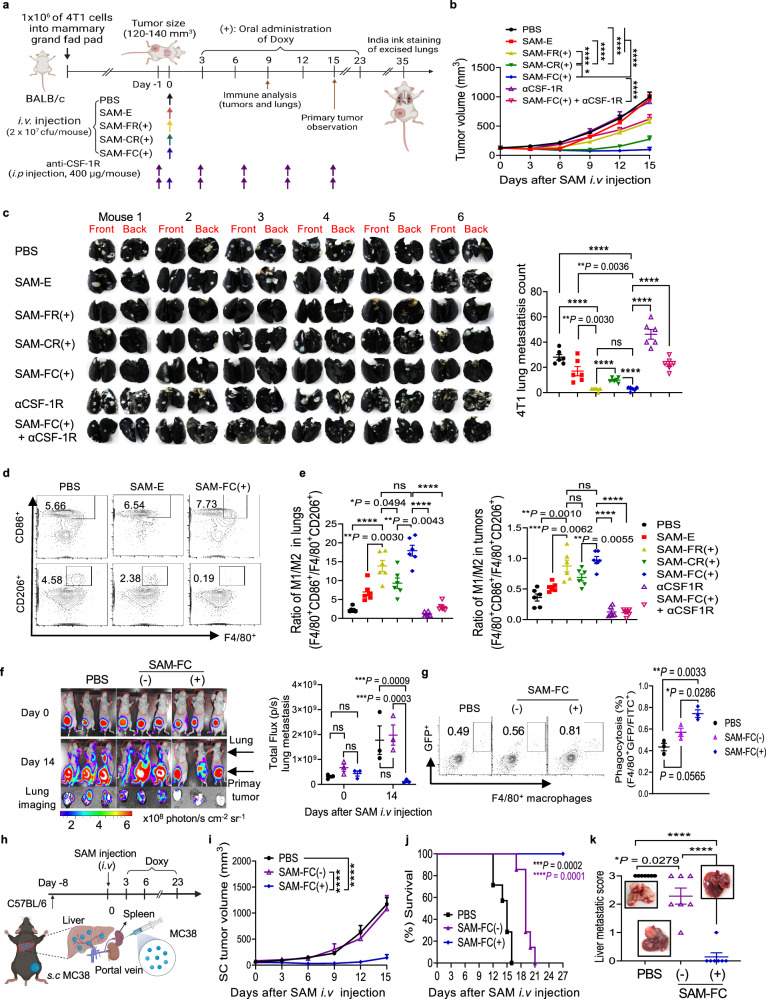
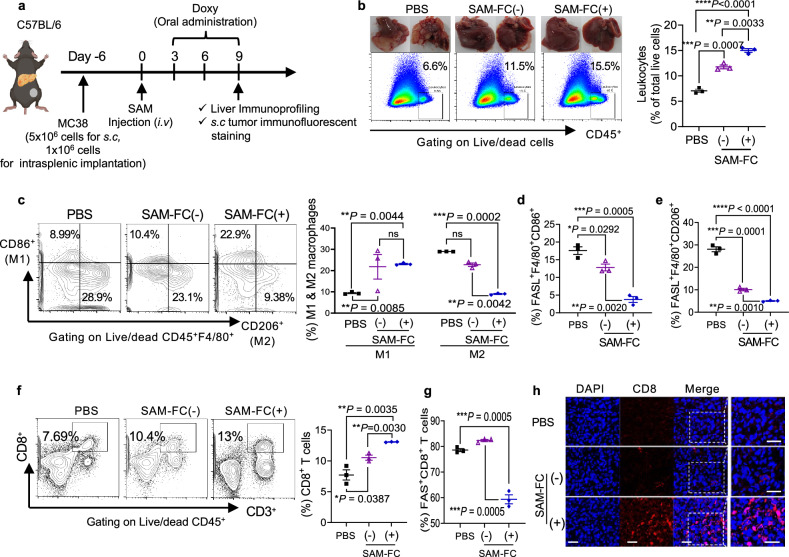
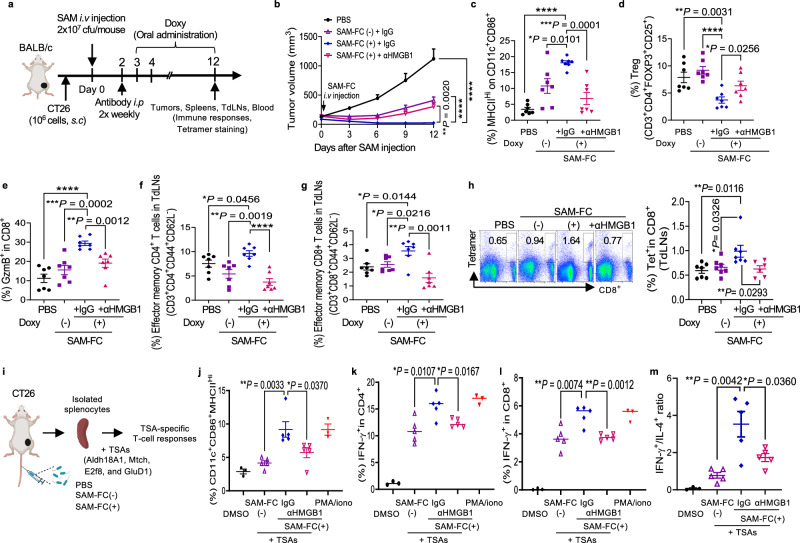
Synergistic combinations of immunotherapeutic agents can improve the performance of anti-cancer therapies but may lead to immune-mediated adverse effects. These side-effects can be overcome by using a tumor-specific delivery system. Here, we report a method of targeted immunotherapy using an attenuated Salmonella typhimurium (SAM-FC) engineered to release dual payloads: cytolysin A (ClyA), a cytolytic anti-cancer agent, and Vibrio vulnificus flagellin B (FlaB), a potent inducer of anti-tumor innate immunity. Localized secretion of ClyA from SAM-FC induces immunogenic cancer cell death and promotes release of tumor-specific antigens and damage-associated molecular patterns, which establish long-term antitumor memory. Localized secretion of FlaB promotes phenotypic and functional remodeling of intratumoral macrophages that markedly inhibits tumor metastasis in mice bearing tumors of mouse and human origin. Both primary and metastatic tumors from bacteria-treated female mice are characterized by massive infiltration of anti-tumorigenic innate immune cells and activated tumor-specific effector/memory T cells; however, the percentage of immunosuppressive cells is low. Here, we show that SAM-FC induces functional reprogramming of the tumor immune microenvironment by activating both the innate and adaptive arms of the immune system and can be used for targeted delivery of multiple immunotherapeutic payloads for the establishment of potent and long-lasting antitumor immunity.
© 2024. The Author(s).
Conflict of interest statement
A patent for a novel DNA construct aimed at the diagnosis and treatment of cancer (KR102252939) has been registered by J.M., Y.H., and S.Y. as the inventors. In addition, a patent application for an innovative anticancer therapy using engineered bacteria expressing flagellin and pore-forming proteins (KR1020210153355) has been submitted by J.M., Y.H., S.Y., and D.N. as its inventors. CNCure Biotech, Inc. possesses exclusive licenses for these intellectual properties. This IP stands to gain financial benefits upon further validation. The remaining authors declare no competing interests.
Figures



References
MeSH terms
Substances
Grants and funding
- NRF-2020M3A9G3080282/National Research Foundation of Korea (NRF)
- No. 2020R1A5A2031185/National Research Foundation of Korea (NRF)
- 2021M3C1C3097637/National Research Foundation of Korea (NRF)
- 2021C300/National Research Foundation of Korea (NRF)
- NRF-2020M3A9G3080330/National Research Foundation of Korea (NRF)
LinkOut - more resources
Full Text Sources
