

Microsatellite instability at U2AF-binding polypyrimidic tract sites perturbs alternative splicing during colorectal cancer initiation
- PMID: 39107855
- PMCID: PMC11304650
- DOI: 10.1186/s13059-024-03340-5
Microsatellite instability at U2AF-binding polypyrimidic tract sites perturbs alternative splicing during colorectal cancer initiation
Abstract
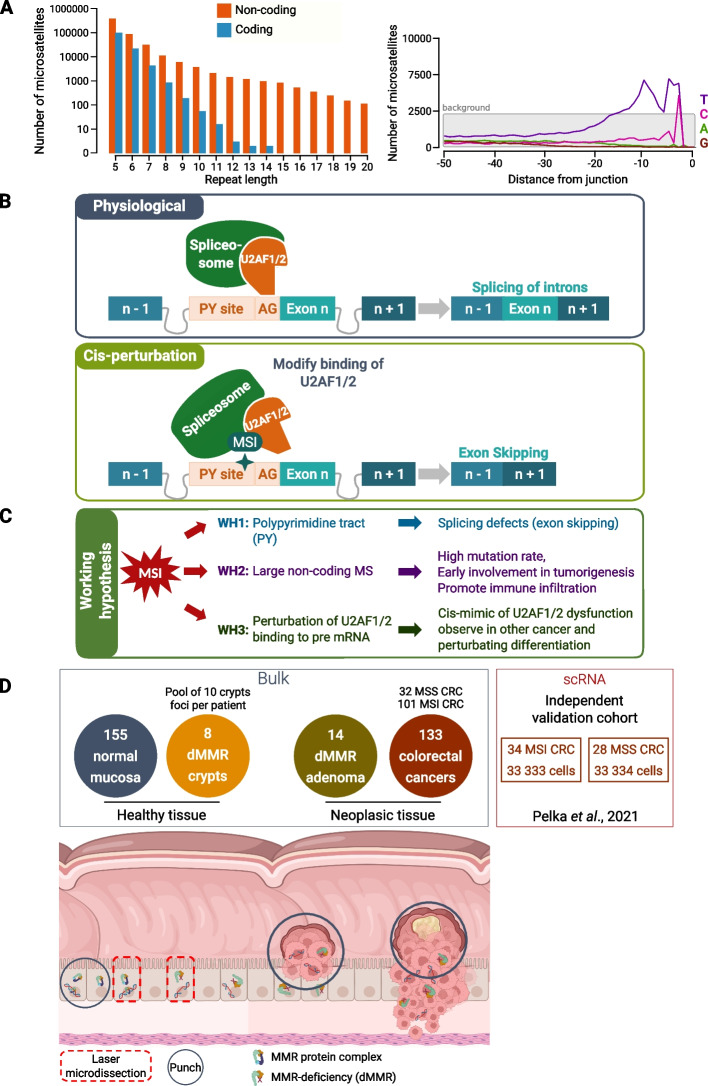
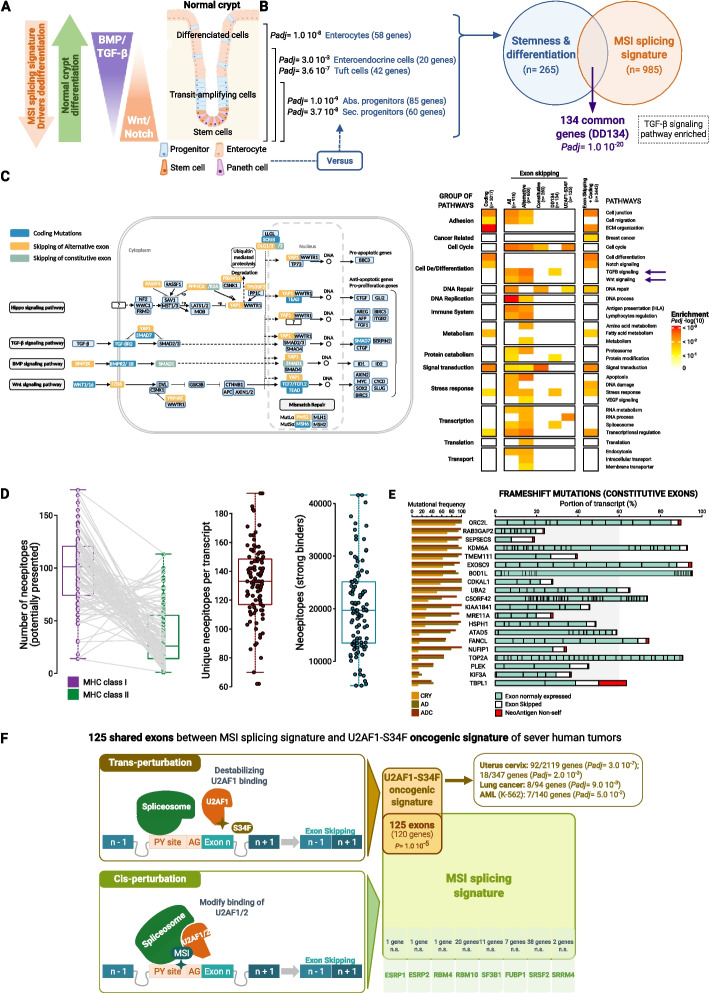
Background: Microsatellite instability (MSI) due to mismatch repair deficiency (dMMR) is common in colorectal cancer (CRC). These cancers are associated with somatic coding events, but the noncoding pathophysiological impact of this genomic instability is yet poorly understood. Here, we perform an analysis of coding and noncoding MSI events at the different steps of colorectal tumorigenesis using whole exome sequencing and search for associated splicing events via RNA sequencing at the bulk-tumor and single-cell levels.
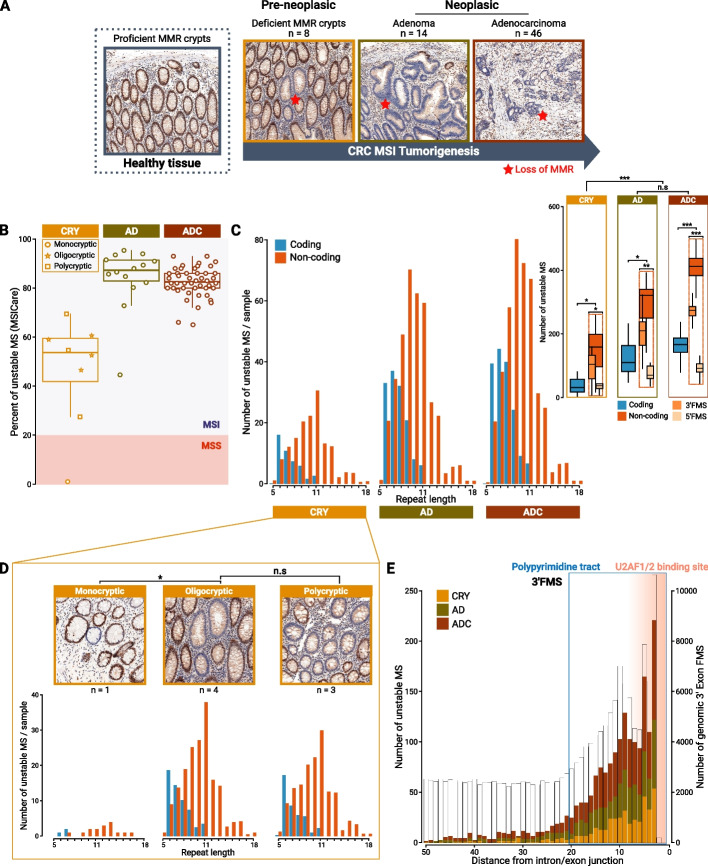
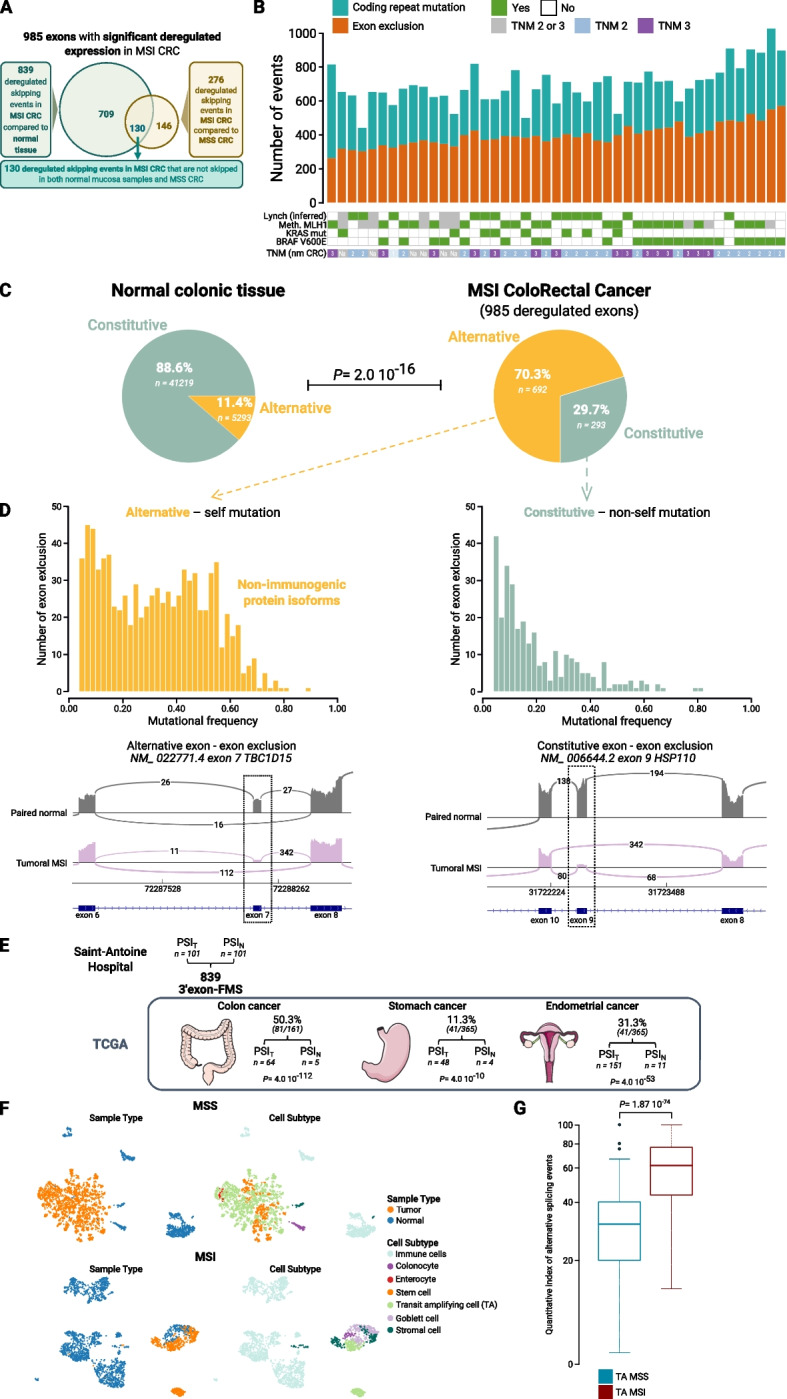
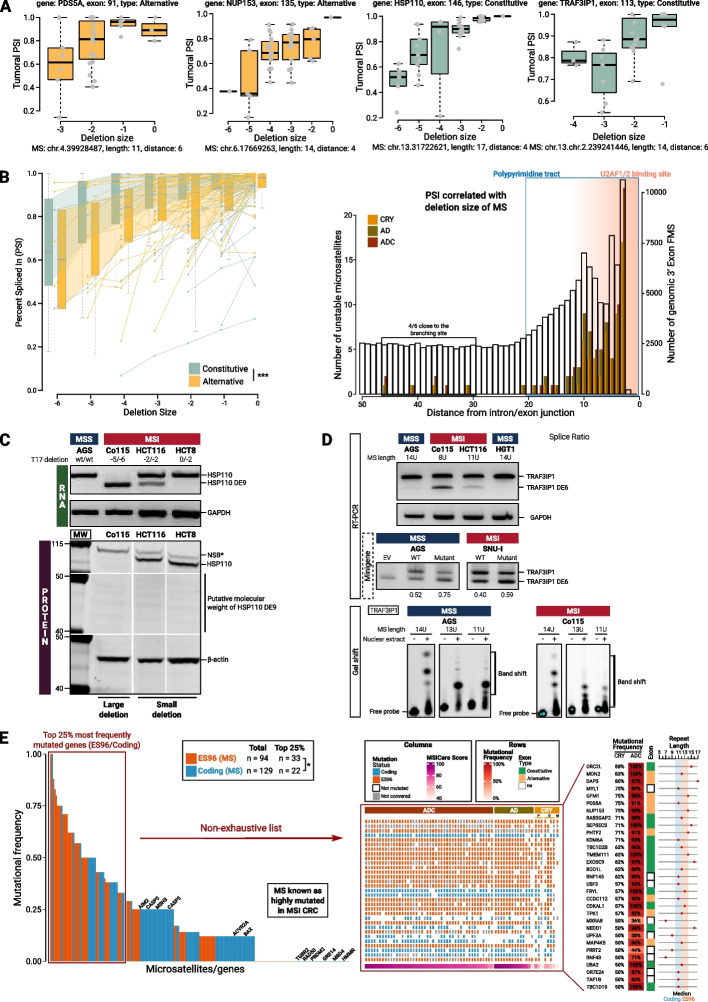
Results: Our results demonstrate that MSI leads to hundreds of noncoding DNA mutations, notably at polypyrimidine U2AF RNA-binding sites which are endowed with cis-activity in splicing, while higher frequency of exon skipping events are observed in the mRNAs of MSI compared to non-MSI CRC. At the DNA level, these noncoding MSI mutations occur very early prior to cell transformation in the dMMR colonic crypt, accounting for only a fraction of the exon skipping in MSI CRC. At the RNA level, the aberrant exon skipping signature is likely to impair colonic cell differentiation in MSI CRC affecting the expression of alternative exons encoding protein isoforms governing cell fate, while also targeting constitutive exons, making dMMR cells immunogenic in early stage before the onset of coding mutations. This signature is characterized by its similarity to the oncogenic U2AF1-S34F splicing mutation observed in several other non-MSI cancer.
Conclusions: Overall, these findings provide evidence that a very early RNA splicing signature partly driven by MSI impairs cell differentiation and promotes MSI CRC initiation, far before coding mutations which accumulate later during MSI tumorigenesis.
Keywords: Alternative splicing; Cancer initiation; Cell differentiation; Colonic crypt; Colorectal cancer; Microsatellite instability; Mismatch repair deficiency; Polypyrimidine U2AF binding site; Whole-exome and RNA sequencing.
© 2024. The Author(s).
Conflict of interest statement
PDLG and AJ (authors) declare a conflict of interest with the Genosplice company.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
