

Role of RNA structural plasticity in modulating HIV-1 genome packaging and translation
- PMID: 39110735
- PMCID: PMC11331132
- DOI: 10.1073/pnas.2407400121
Role of RNA structural plasticity in modulating HIV-1 genome packaging and translation
Abstract
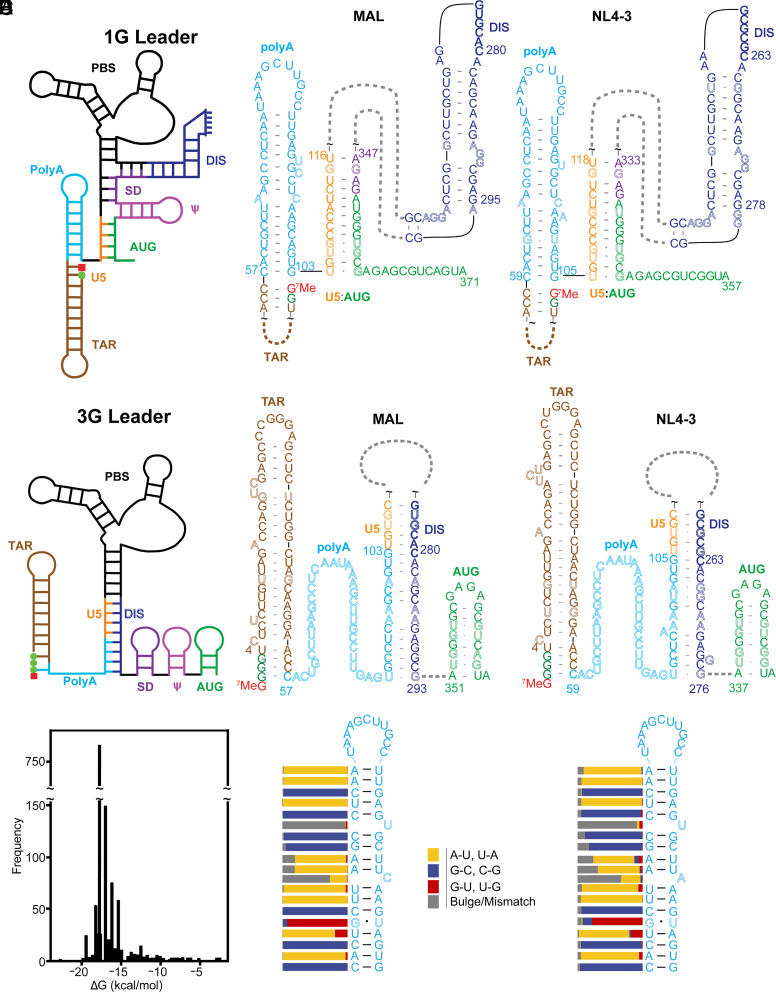
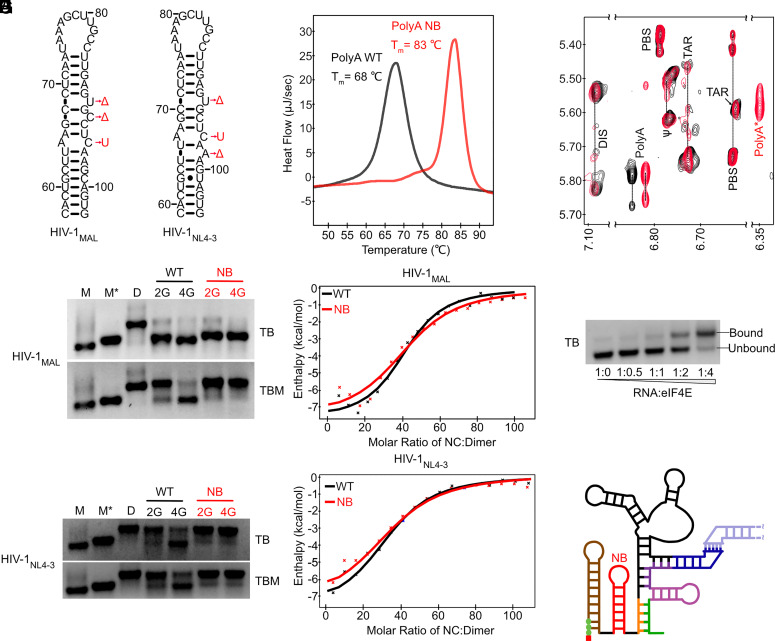
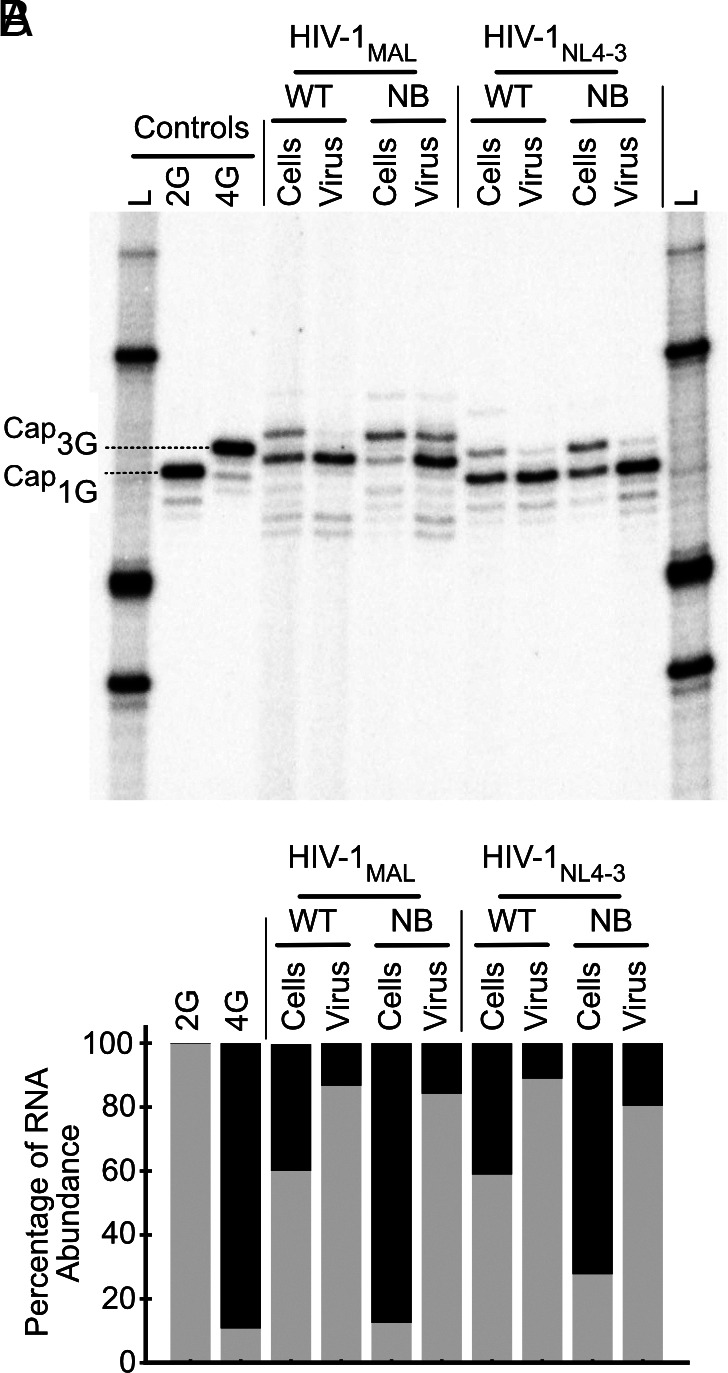
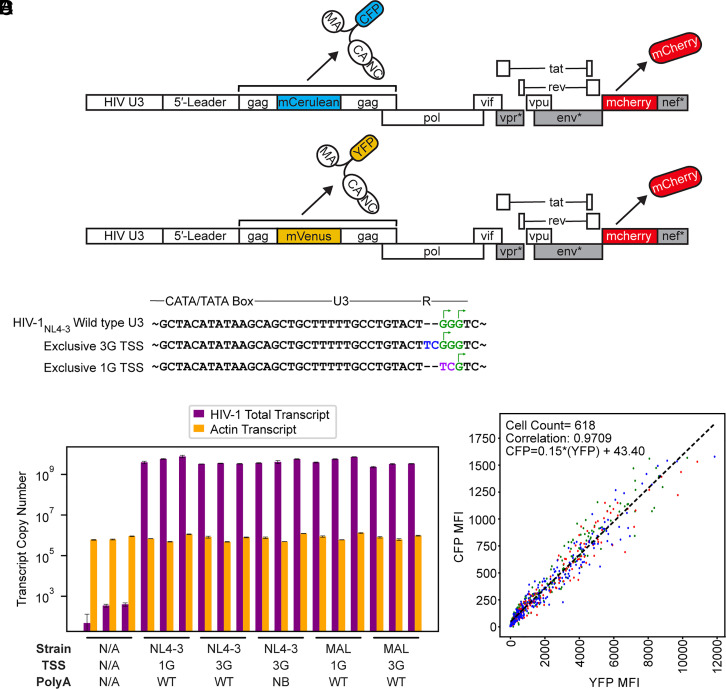
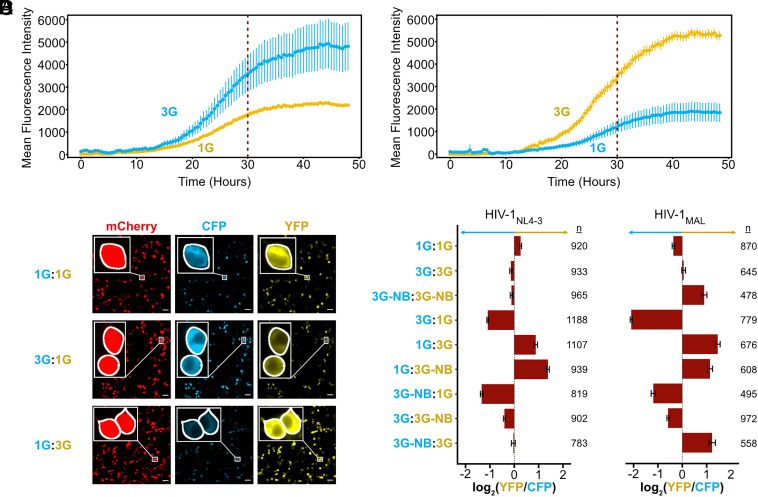
HIV-1 transcript function is controlled in part by twinned transcriptional start site usage, where 5' capped RNAs beginning with a single guanosine (1G) are preferentially packaged into progeny virions as genomic RNA (gRNA) whereas those beginning with three sequential guanosines (3G) are retained in cells as mRNAs. In 3G transcripts, one of the additional guanosines base pairs with a cytosine located within a conserved 5' polyA element, resulting in formation of an extended 5' polyA structure as opposed to the hairpin structure formed in 1G RNAs. To understand how this remodeling influences overall transcript function, we applied in vitro biophysical studies with in-cell genome packaging and competitive translation assays to native and 5' polyA mutant transcripts generated with promoters that differentially produce 1G or 3G RNAs. We identified mutations that stabilize the 5' polyA hairpin structure in 3G RNAs, which promote RNA dimerization and Gag binding without sequestering the 5' cap. None of these 3G transcripts were competitively packaged, confirming that cap exposure is a dominant negative determinant of viral genome packaging. For all RNAs examined, conformations that favored 5' cap exposure were both poorly packaged and more efficiently translated than those that favored 5' cap sequestration. We propose that structural plasticity of 5' polyA and other conserved RNA elements place the 5' leader on a thermodynamic tipping point for low-energetic (~3 kcal/mol) control of global transcript structure and function.
Keywords: HIV-1; dimerization; packaging; polyA; translation.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures
References
-
- Coffin J. M., Hughes S. H., Varmus H. E., Retroviruses (Cold Spring Harbor Laboratory Press, Plainview, N.Y., 1997). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
