

Genomic data provide insights into the classification of extant termites
- PMID: 39112457
- PMCID: PMC11306793
- DOI: 10.1038/s41467-024-51028-y
Genomic data provide insights into the classification of extant termites
Abstract
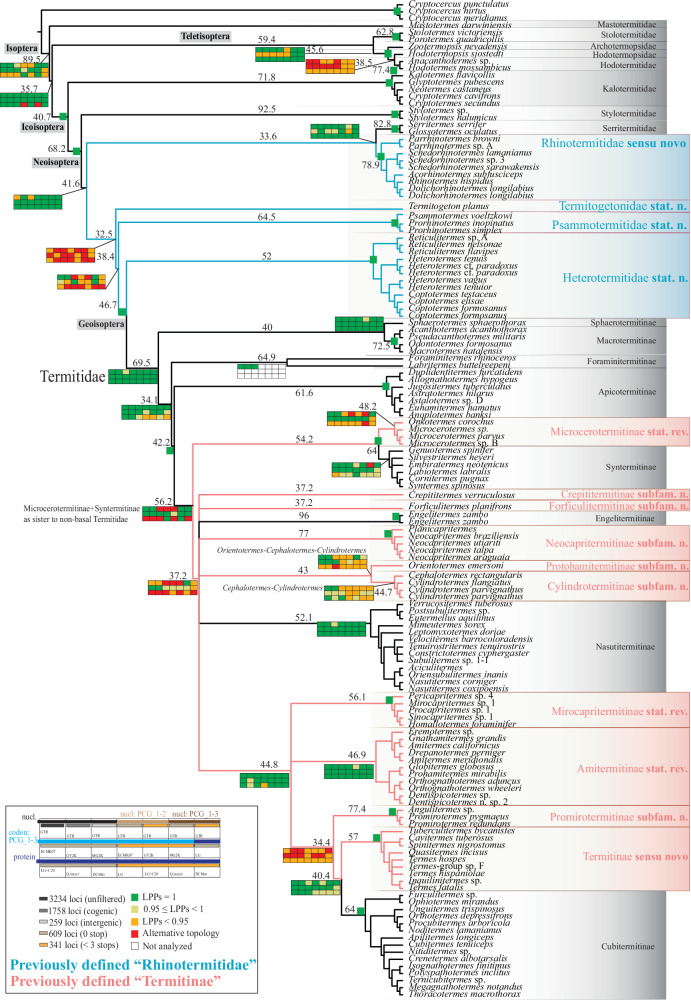
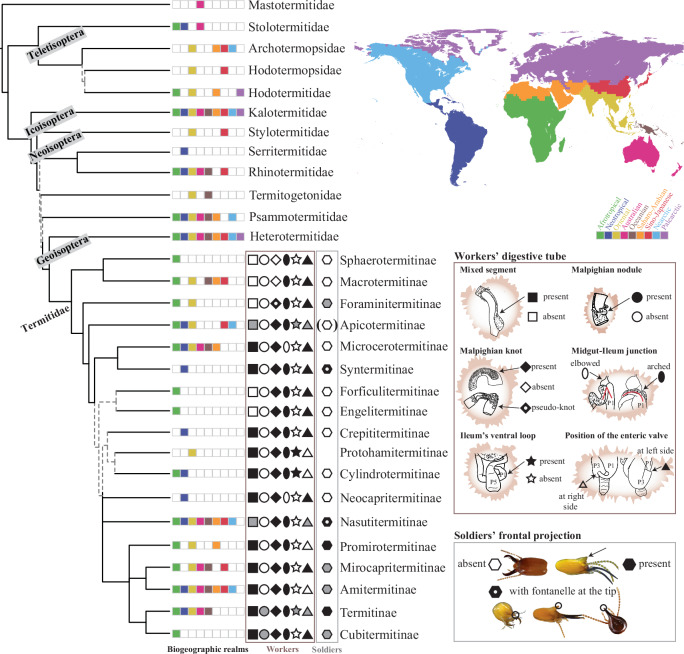
The higher classification of termites requires substantial revision as the Neoisoptera, the most diverse termite lineage, comprise many paraphyletic and polyphyletic higher taxa. Here, we produce an updated termite classification using genomic-scale analyses. We reconstruct phylogenies under diverse substitution models with ultraconserved elements analyzed as concatenated matrices or within the multi-species coalescence framework. Our classification is further supported by analyses controlling for rogue loci and taxa, and topological tests. We show that the Neoisoptera are composed of seven family-level monophyletic lineages, including the Heterotermitidae Froggatt, Psammotermitidae Holmgren, and Termitogetonidae Holmgren, raised from subfamilial rank. The species-rich Termitidae are composed of 18 subfamily-level monophyletic lineages, including the new subfamilies Crepititermitinae, Cylindrotermitinae, Forficulitermitinae, Neocapritermitinae, Protohamitermitinae, and Promirotermitinae; and the revived Amitermitinae Kemner, Microcerotermitinae Holmgren, and Mirocapritermitinae Kemner. Building an updated taxonomic classification on the foundation of unambiguously supported monophyletic lineages makes it highly resilient to potential destabilization caused by the future availability of novel phylogenetic markers and methods. The taxonomic stability is further guaranteed by the modularity of the new termite classification, designed to accommodate as-yet undescribed species with uncertain affinities to the herein delimited monophyletic lineages in the form of new families or subfamilies.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Grimaldi, D. A. & Engel, M. S. Evolution of the Insects. (Cambridge University Press, 2005).
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
