

Age-related epithelial defects limit thymic function and regeneration
- PMID: 39112630
- PMCID: PMC11362016
- DOI: 10.1038/s41590-024-01915-9
Age-related epithelial defects limit thymic function and regeneration
Abstract
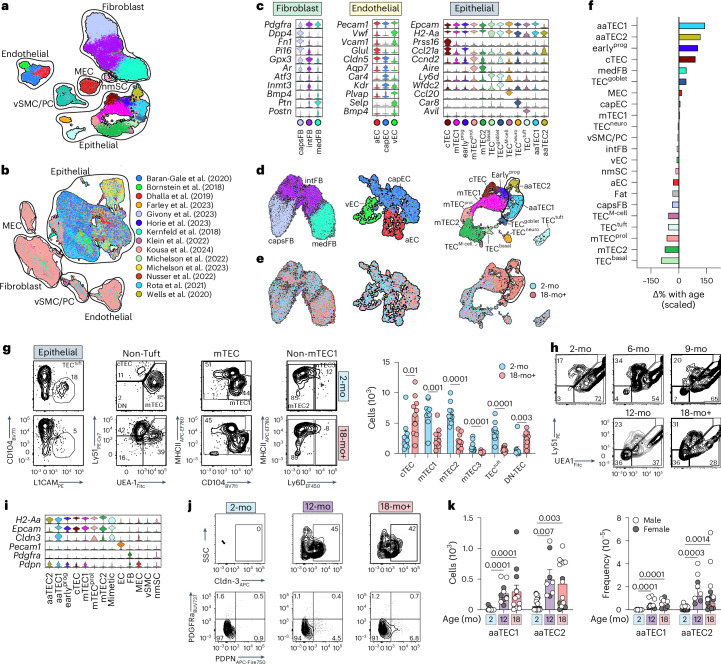
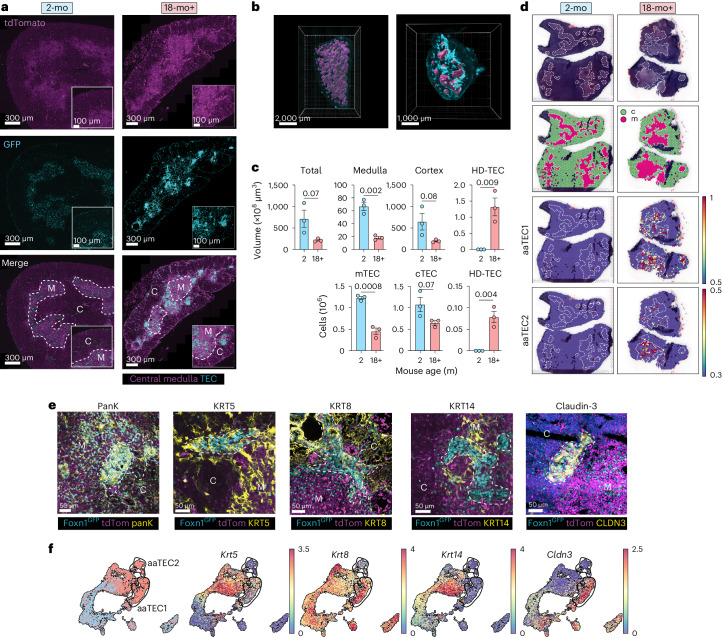
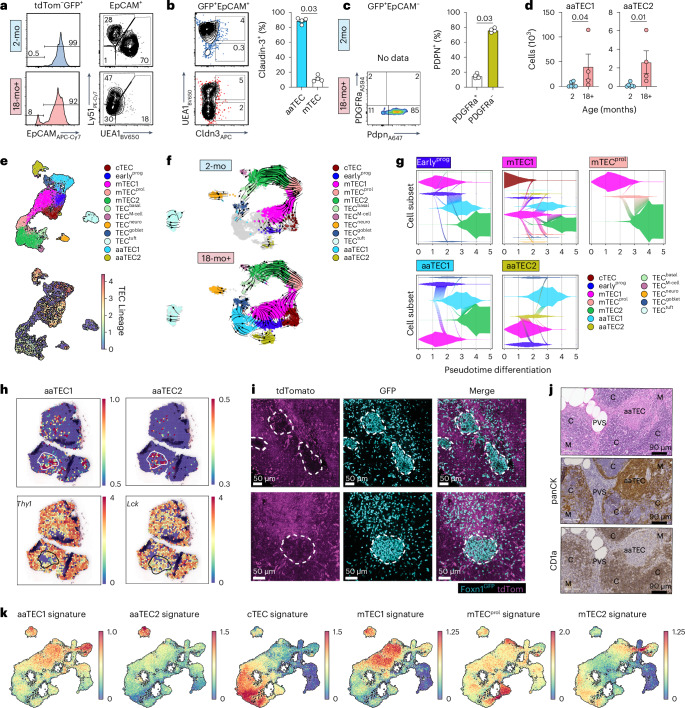
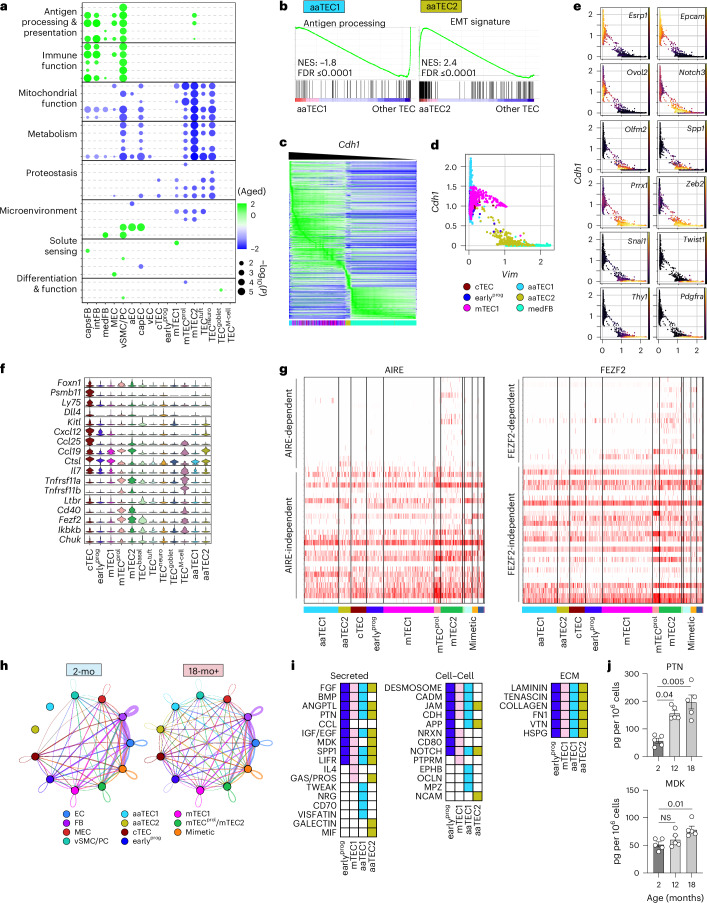
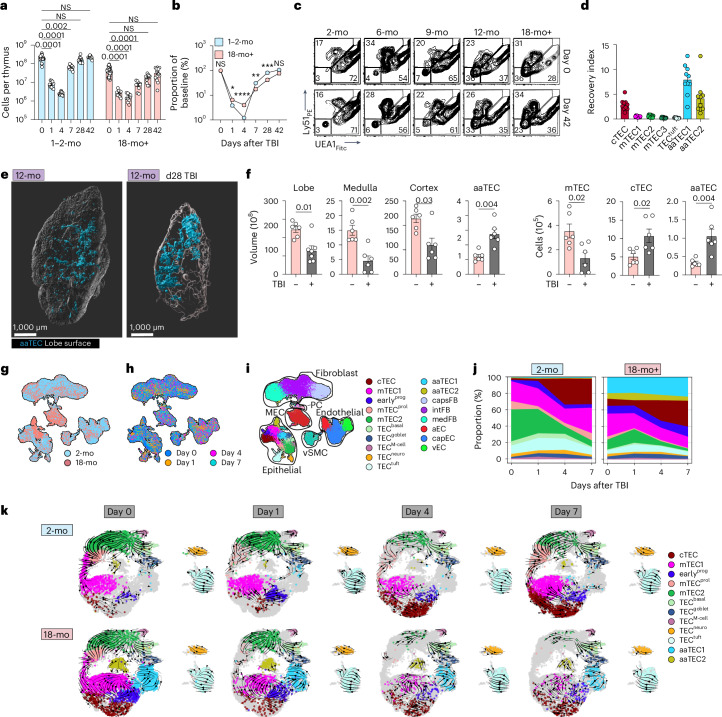
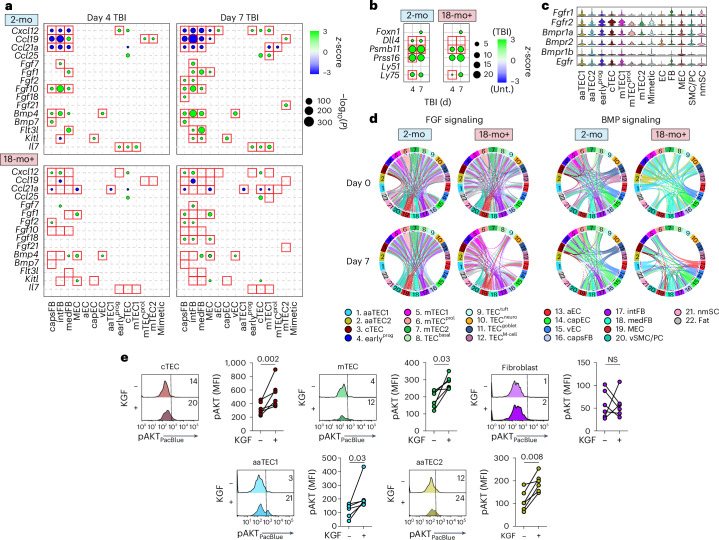
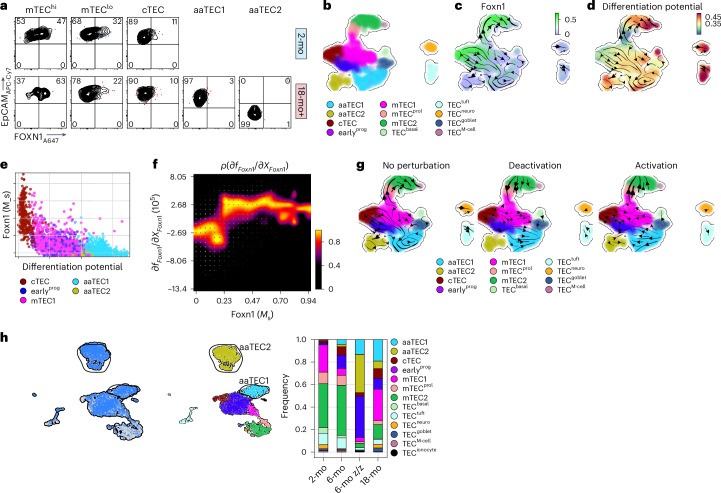
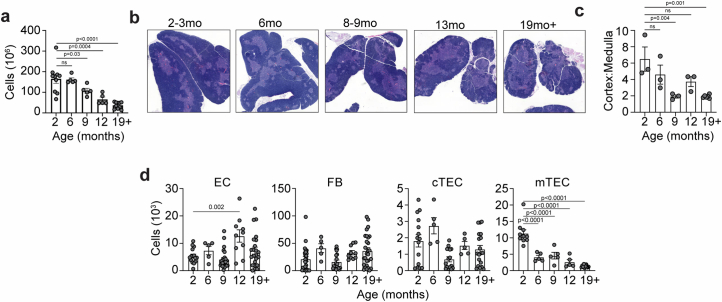
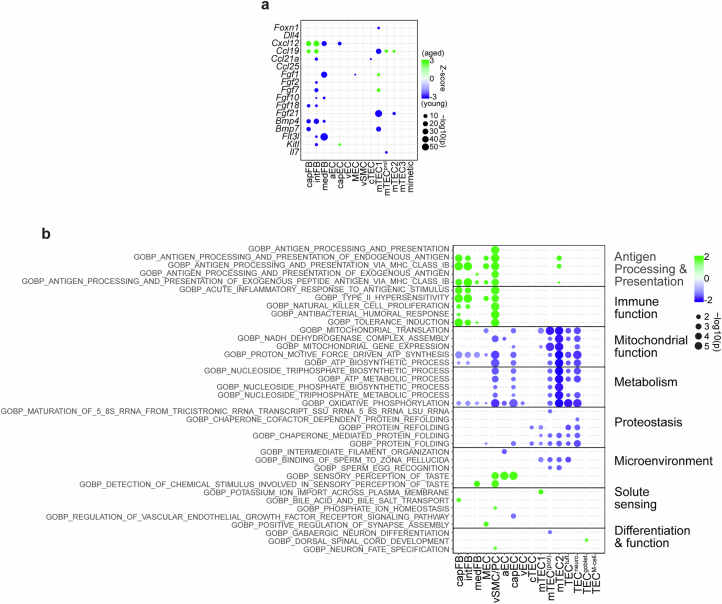
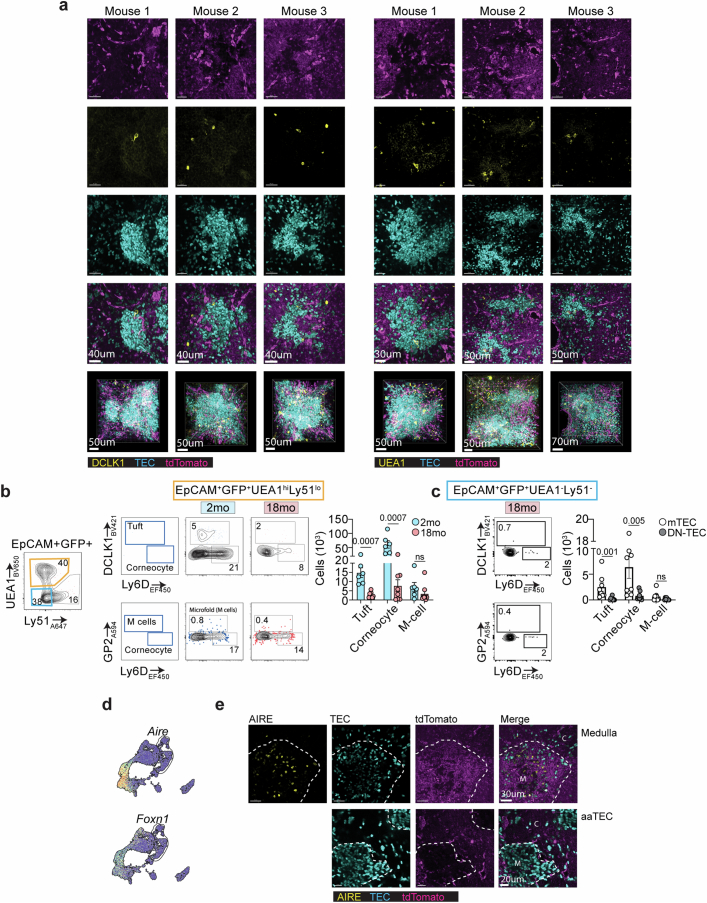
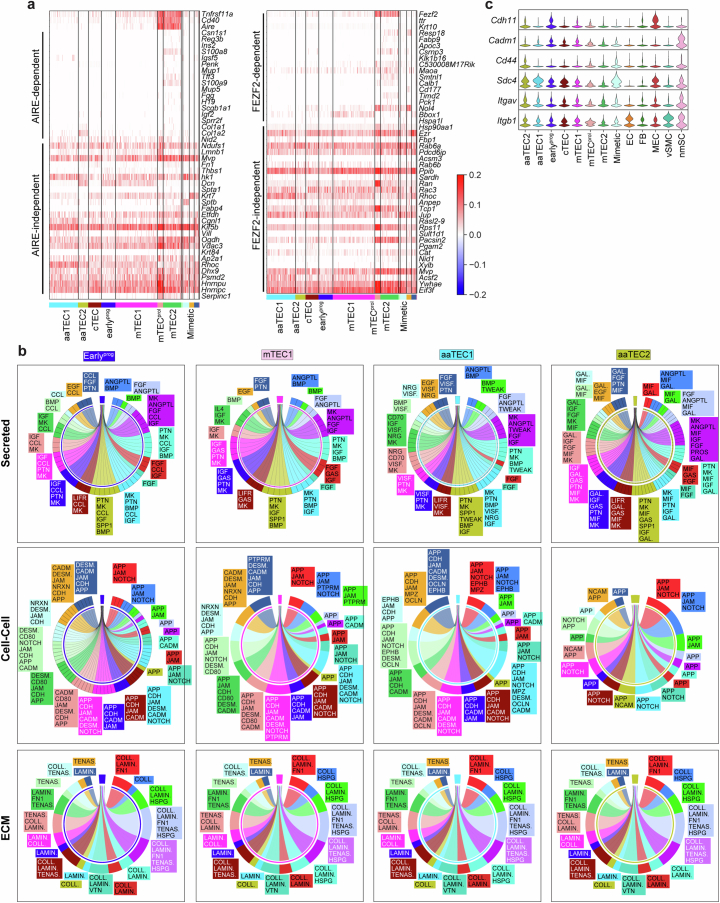
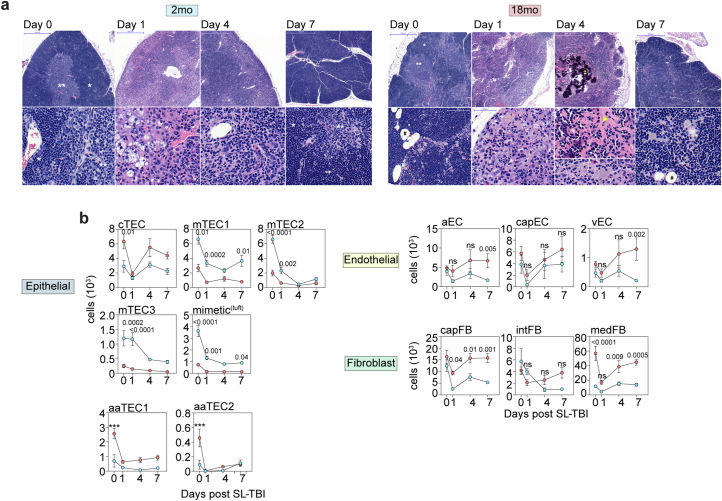
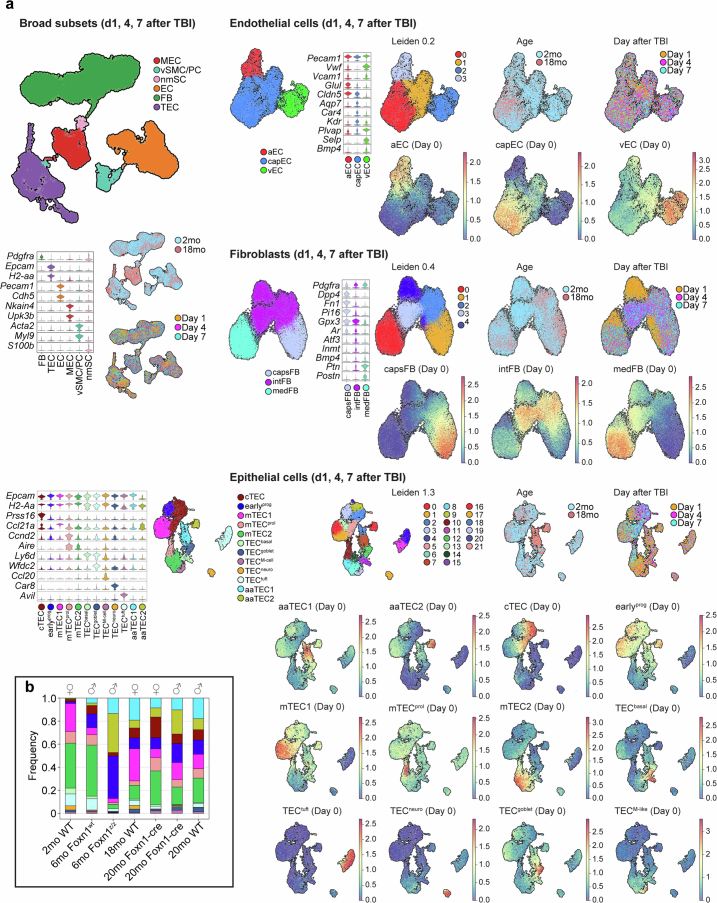
The thymus is essential for establishing adaptive immunity yet undergoes age-related involution that leads to compromised immune responsiveness. The thymus is also extremely sensitive to acute insult and although capable of regeneration, this capacity declines with age for unknown reasons. We applied single-cell and spatial transcriptomics, lineage-tracing and advanced imaging to define age-related changes in nonhematopoietic stromal cells and discovered the emergence of two atypical thymic epithelial cell (TEC) states. These age-associated TECs (aaTECs) formed high-density peri-medullary epithelial clusters that were devoid of thymocytes; an accretion of nonproductive thymic tissue that worsened with age, exhibited features of epithelial-to-mesenchymal transition and was associated with downregulation of FOXN1. Interaction analysis revealed that the emergence of aaTECs drew tonic signals from other functional TEC populations at baseline acting as a sink for TEC growth factors. Following acute injury, aaTECs expanded substantially, further perturbing trophic regeneration pathways and correlating with defective repair of the involuted thymus. These findings therefore define a unique feature of thymic involution linked to immune aging and could have implications for developing immune-boosting therapies in older individuals.
© 2024. The Author(s).
Conflict of interest statement
M.R.M.vdB. has received research support and stock options from Seres Therapeutics and stock options from Notch Therapeutics and Pluto Therapeutics; has consulted, received honorarium from or participated in advisory boards for Seres Therapeutics, WindMIL Therapeutics, Rheos Medicines, Merck & Co, Magenta Therapeutics, Frazier Healthcare Partners, Nektar Therapeutics, Notch Therapeutics, Forty Seven, Priothera, Ceramedix, Lygenesis, Pluto Therapeutics, GlaskoSmithKline, Da Volterra, Vor BioPharma, Novartis (Spouse), Synthekine (Spouse) and Beigene (Spouse); he has IP Licensing with Seres Therapeutics and Juno Therapeutics; and holds a fiduciary role on the Foundation Board of DKMS (a nonprofit organization). J.A.D. and M.R.M.vdB. are founders of, and receive stock options, from ThymoFox; and both have received royalties from Wolters Kluwer. The Walter and Eliza Hall Institute of Medical Research receives milestone and royalty payments related to venetoclax. Employees are entitled to receive benefits related to these payments; D.H.D.G. reports receiving benefits. D.H.D.G. has received research funding from Servier. The remaining authors declare no competing interests.
Figures
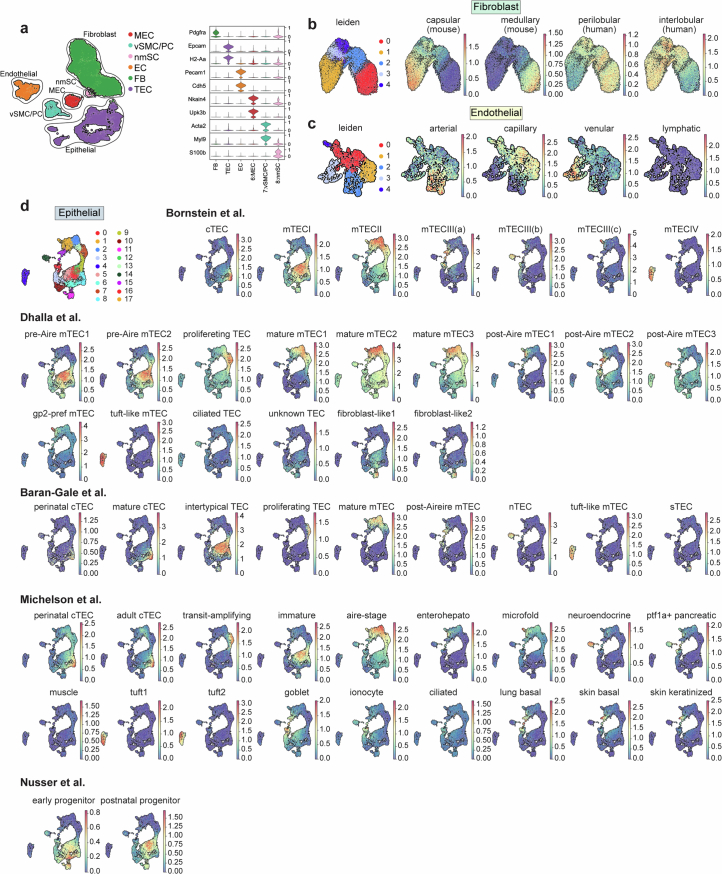
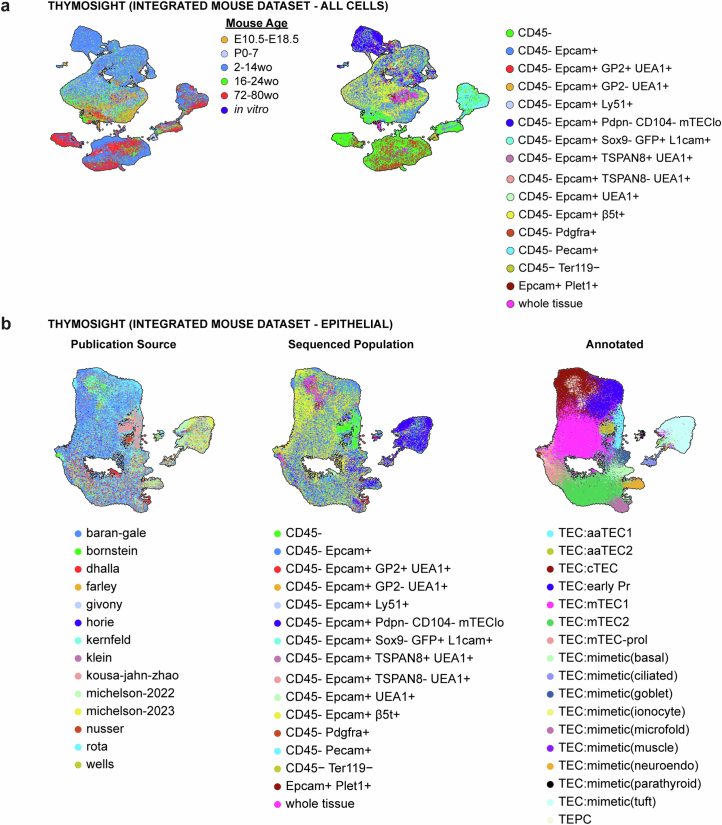
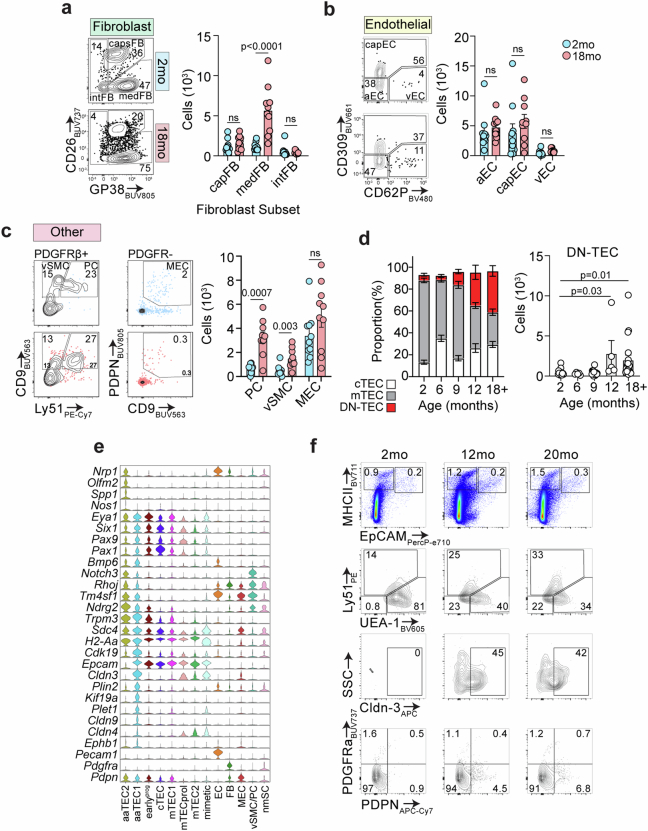
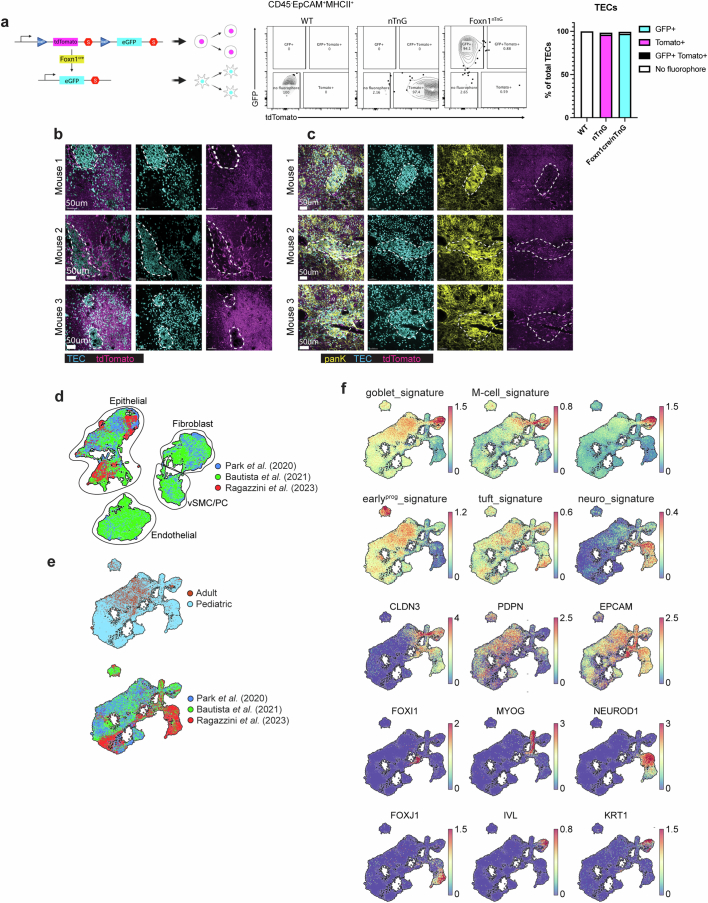
References
MeSH terms
Substances
Grants and funding
- T32-GM007270/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- 1187367/Department of Health | National Health and Medical Research Council (NHMRC)
- R01 CA228308/CA/NCI NIH HHS/United States
- 1158024/Department of Health | National Health and Medical Research Council (NHMRC)
- R01-HL145276/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- R01-HL147584/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- R01 HL123340/HL/NHLBI NIH HHS/United States
- R01 HL145276/HL/NHLBI NIH HHS/United States
- R01-CA228308/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
- T32 GM007103/GM/NIGMS NIH HHS/United States
- P30 CA015704/CA/NCI NIH HHS/United States
- P01 CA023766/CA/NCI NIH HHS/United States
- R01 HL165673/HL/NHLBI NIH HHS/United States
- R01 HL147584/HL/NHLBI NIH HHS/United States
- P01-AG052359/U.S. Department of Health & Human Services | NIH | National Institute on Aging (U.S. National Institute on Aging)
- P30-CA015704/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
- K08 CA293271/CA/NCI NIH HHS/United States
- 1090236/Department of Health | National Health and Medical Research Council (NHMRC)
- P30 CA008748/CA/NCI NIH HHS/United States
- P01 AG052359/AG/NIA NIH HHS/United States
- T32 GM007270/GM/NIGMS NIH HHS/United States
- 1102104/Cancer Council Victoria
- 1078763/Department of Health | National Health and Medical Research Council (NHMRC)
- 1146518/Cancer Council Victoria
- U01-AI70035/U.S. Department of Health & Human Services | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- R35 HL171556/HL/NHLBI NIH HHS/United States
- ALTF-431-2017/European Molecular Biology Organization (EMBO)
- R01 CA228358/CA/NCI NIH HHS/United States
- F30-HL165761/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- R01-HL123340/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- R35-HL-171556/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- 1121325/Department of Health | National Health and Medical Research Council (NHMRC)
- F30 HL165761/HL/NHLBI NIH HHS/United States
- U01 AI170035/AI/NIAID NIH HHS/United States
- R01-HL165673/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
