

Seed longevity is controlled by metacaspases
- PMID: 39117606
- PMCID: PMC11310522
- DOI: 10.1038/s41467-024-50848-2
Seed longevity is controlled by metacaspases
Abstract
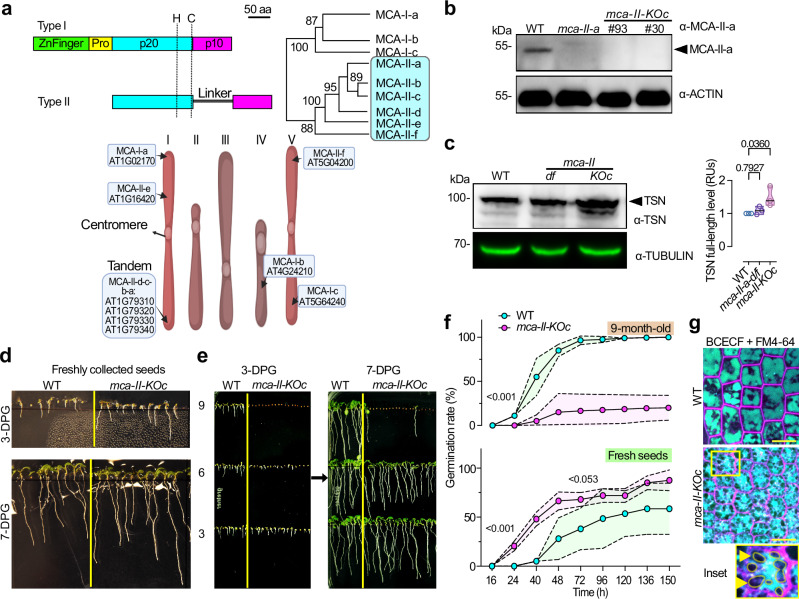
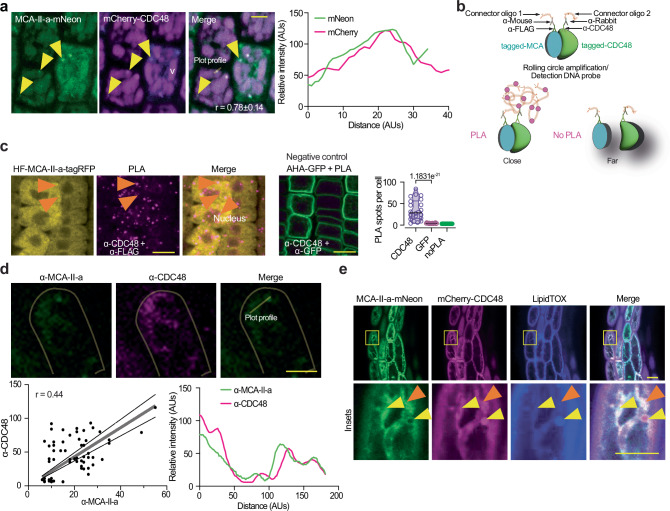
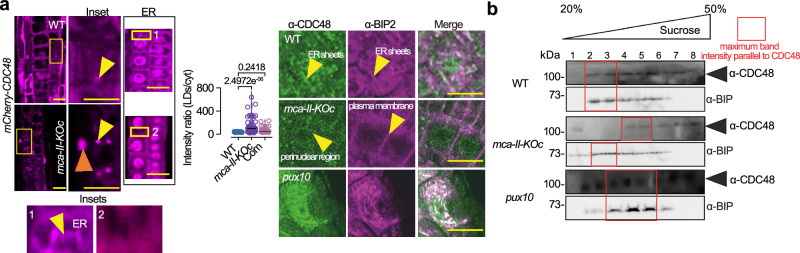
To survive extreme desiccation, seeds enter a period of quiescence that can last millennia. Seed quiescence involves the accumulation of protective storage proteins and lipids through unknown adjustments in protein homeostasis (proteostasis). Here, we show that mutation of all six type-II metacaspase (MCA-II) proteases in Arabidopsis thaliana disturbs proteostasis in seeds. MCA-II mutant seeds fail to restrict the AAA ATPase CELL DIVISION CYCLE 48 (CDC48) at the endoplasmic reticulum to discard misfolded proteins, compromising seed storability. Endoplasmic reticulum (ER) localization of CDC48 relies on the MCA-IIs-dependent cleavage of PUX10 (ubiquitination regulatory X domain-containing 10), the adaptor protein responsible for titrating CDC48 to lipid droplets. PUX10 cleavage enables the shuttling of CDC48 between lipid droplets and the ER, providing an important regulatory mechanism sustaining spatiotemporal proteolysis, lipid droplet dynamics, and protein homeostasis. In turn, the removal of the PUX10 adaptor in MCA-II mutant seeds partially restores proteostasis, CDC48 localization, and lipid droplet dynamics prolonging seed lifespan. Taken together, we uncover a proteolytic module conferring seed longevity.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures







References
MeSH terms
Substances
Grants and funding
- 2018.0026/Knut och Alice Wallenbergs Stiftelse (Knut and Alice Wallenberg Foundation)
- 2018.0026/Knut och Alice Wallenbergs Stiftelse (Knut and Alice Wallenberg Foundation)
- 101039824/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 European Research Council (H2020 Excellent Science - European Research Council)
- 101126019/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 European Research Council (H2020 Excellent Science - European Research Council)
- BB/R012172/1/RCUK | Biotechnology and Biological Sciences Research Council (BBSRC)
- 872969/EC | Horizon 2020 Framework Programme (EU Framework Programme for Research and Innovation H2020)
- 2019-04250/Vetenskapsrådet (Swedish Research Council)
- 2019-04250/Vetenskapsrådet (Swedish Research Council)
- 656011/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 Marie Skłodowska-Curie Actions (H2020 Excellent Science - Marie Skłodowska-Curie Actions)
- 22:2025/Carl Tryggers Stiftelse för Vetenskaplig Forskning (Carl Trygger Foundation)
LinkOut - more resources
Full Text Sources
