

Hepatocyte-specific loss of melanocortin 1 receptor disturbs fatty acid metabolism and promotes adipocyte hypertrophy
- PMID: 39117851
- PMCID: PMC11502480
- DOI: 10.1038/s41366-024-01600-9
Hepatocyte-specific loss of melanocortin 1 receptor disturbs fatty acid metabolism and promotes adipocyte hypertrophy
Abstract
Background/objectives: Melanocortins mediate their biological functions via five different melanocortin receptors (MC1R - MC5R). MC1R is expressed in the skin and leukocytes, where it regulates skin pigmentation and inflammatory responses. MC1R is also present in the liver and white adipose tissue, but its functional role in these tissues is unclear. This study aimed at determining the regulatory role of MC1R in fatty acid metabolism.
Methods: Male recessive yellow (Mc1re/e) mice, a model of global MC1R deficiency, and male hepatocyte-specific MC1R deficient mice (Mc1r LKO) were fed a chow or Western diet for 12 weeks. The mouse models were characterized for body weight and composition, liver adiposity, adipose tissue mass and morphology, glucose metabolism and lipid metabolism. Furthermore, qPCR and RNA sequencing analyses were used to investigate gene expression profiles in the liver and adipose tissue. HepG2 cells and primary mouse hepatocytes were used to study the effects of pharmacological MC1R activation.
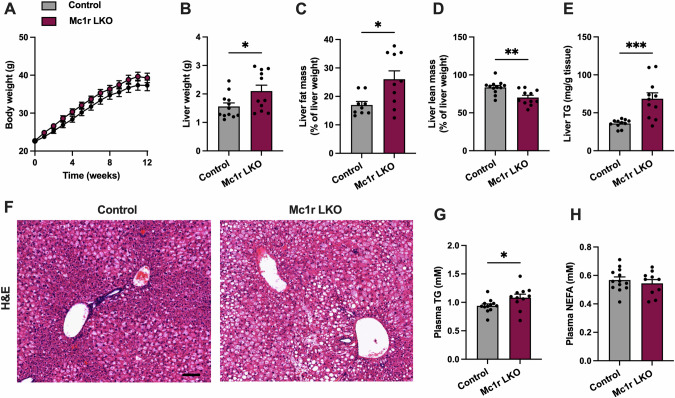
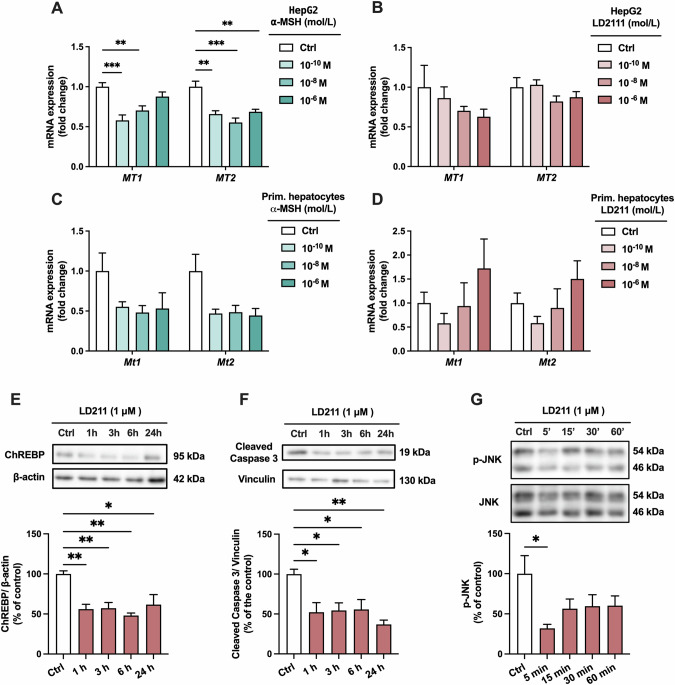
Results: Chow- and Western diet-fed Mc1re/e showed increased liver weight, white adipose tissue mass and plasma triglyceride (TG) concentration compared to wild type mice. This phenotype occurred without significant changes in food intake, body weight, physical activity or glucose metabolism. Mc1r LKO mice displayed a similar phenotype characterized by larger fat depots, increased adipocyte hypertrophy and enhanced accumulation of TG in the liver and plasma. In terms of gene expression, markers of de novo lipogenesis, inflammation and apoptosis were upregulated in the liver of Mc1r LKO mice, while enzymes regulating lipolysis were downregulated in white adipose tissue of these mice. In cultured hepatocytes, selective activation of MC1R reduced ChREBP expression, which is a central transcription factor for lipogenesis.
Conclusions: Hepatocyte-specific loss of MC1R disturbs fatty acid metabolism in the liver and leads to an obesity phenotype characterized by enhanced adipocyte hypertrophy and TG accumulation in the liver and circulation.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Eckel RH, Grundy SM, Zimmet PZ. The metabolic syndrome. Lancet. 2005;365:1415–28. - PubMed
-
- Bugianesi E, Gastaldelli A, Vanni E, Gambino R, Cassader M, Baldi S, et al. Insulin resistance in non-diabetic patients with non-alcoholic fatty liver disease: sites and mechanisms. Diabetologia. 2005;48:634–42. - PubMed
-
- Haas JT, Francque S, Staels B. Pathophysiology and mechanisms of nonalcoholic fatty liver disease. Annu Rev Physiol. 2016;78:181–205. - PubMed
-
- Azzu V, Vacca M, Virtue S, Allison M, Vidal-Puig A. Adipose tissue-liver cross talk in the control of whole-body metabolism: implications in nonalcoholic fatty liver disease. Gastroenterology. 2020;158:1899–912. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
