

Blocking HXA3-mediated neutrophil elastase release during S. pneumoniae lung infection limits pulmonary epithelial barrier disruption and bacteremia
- PMID: 39120139
- PMCID: PMC11389395
- DOI: 10.1128/mbio.01856-24
Blocking HXA3-mediated neutrophil elastase release during S. pneumoniae lung infection limits pulmonary epithelial barrier disruption and bacteremia
Abstract
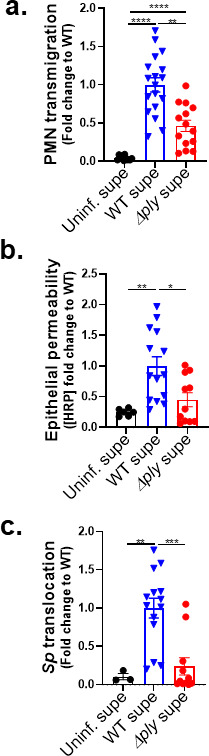
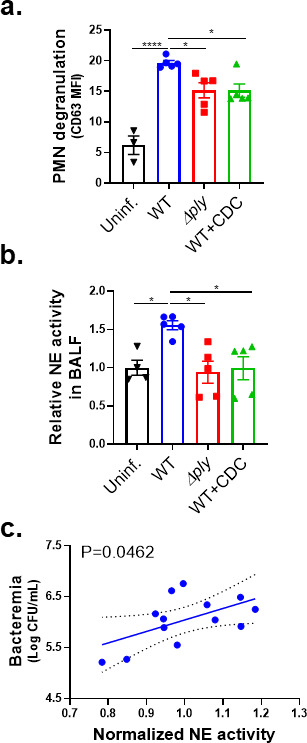
Streptococcus pneumoniae (Sp), a leading cause of community-acquired pneumonia, can spread from the lung into the bloodstream to cause septicemia and meningitis, with a concomitant threefold increase in mortality. Limitations in vaccine efficacy and a rise in antimicrobial resistance have spurred searches for host-directed therapies that target pathogenic immune processes. Polymorphonuclear leukocytes (PMNs) are essential for infection control but can also promote tissue damage and pathogen spread. The major Sp virulence factor, pneumolysin, triggers acute inflammation by stimulating the 12-lipoxygenase (12-LOX) eicosanoid synthesis pathway in epithelial cells. This pathway is required for systemic spread in a mouse pneumonia model and produces a number of bioactive lipids, including hepoxilin A3 (HXA3), a hydroxy epoxide PMN chemoattractant that has been hypothesized to facilitate breach of mucosal barriers. To understand how 12-LOX-dependent inflammation promotes dissemination during Sp lung infection and dissemination, we utilized bronchial stem cell-derived air-liquid interface cultures that lack this enzyme to show that HXA3 methyl ester (HXA3-ME) is sufficient to promote basolateral-to-apical PMN transmigration, monolayer disruption, and concomitant Sp barrier breach. In contrast, PMN transmigration in response to the non-eicosanoid chemoattractant N-formyl-L-methionyl-L-leucyl-phenylalanine (fMLP) did not lead to epithelial disruption or bacterial translocation. Correspondingly, HXA3-ME but not fMLP increased the release of neutrophil elastase (NE) from Sp-infected PMNs. Pharmacologic blockade of NE secretion or activity diminished epithelial barrier disruption and bacteremia after pulmonary challenge of mice. Thus, HXA3 promotes barrier-disrupting PMN transmigration and NE release, pathological events that can be targeted to curtail systemic disease following pneumococcal pneumonia.IMPORTANCEStreptococcus pneumoniae (Sp), a leading cause of pneumonia, can spread from the lung into the bloodstream to cause systemic disease. Limitations in vaccine efficacy and a rise in antimicrobial resistance have spurred searches for host-directed therapies that limit pathologic host immune responses to Sp. Excessive polymorphonuclear leukocyte (PMN) infiltration into Sp-infected airways promotes systemic disease. Using stem cell-derived respiratory cultures that reflect bona fide lung epithelium, we identified eicosanoid hepoxilin A3 as a critical pulmonary PMN chemoattractant that is sufficient to drive PMN-mediated epithelial damage by inducing the release of neutrophil elastase. Inhibition of the release or activity of this protease in mice limited epithelial barrier disruption and bacterial dissemination, suggesting a new host-directed treatment for Sp lung infection.
Keywords: 12-lipoxygenase; Streptococcus pneumoniae; airway mucosal barrier; neutrophil elastase; neutrophil transmigration.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





Update of
-
Blocking HXA3-mediated neutrophil elastase release during S. pneumoniae lung infection limits pulmonary epithelial barrier disruption and bacteremia.bioRxiv [Preprint]. 2024 Jun 25:2024.06.25.600637. doi: 10.1101/2024.06.25.600637. bioRxiv. 2024. Update in: mBio. 2024 Sep 11;15(9):e0185624. doi: 10.1128/mbio.01856-24. PMID: 38979170 Free PMC article. Updated. Preprint.
References
-
- Backhaus E, Berg S, Andersson R, Ockborn G, Malmström P, Dahl M, Nasic S, Trollfors B. 2016. Epidemiology of invasive pneumococcal infections: manifestations, incidence and case fatality rate correlated to age, gender and risk factors. BMC Infect Dis 16:367. doi:10.1186/s12879-016-1648-2 - DOI - PMC - PubMed
-
- Zumla A, Rao M, Wallis RS, Kaufmann SHE, Rustomjee R, Mwaba P, Vilaplana C, Yeboah-Manu D, Chakaya J, Ippolito G, Azhar E, Hoelscher M, Maeurer M, Host-Directed Therapies Network consortium . 2016. Host-directed therapies for infectious diseases: current status, recent progress, and future prospects. Lancet Infect Dis 16:e47–63. doi:10.1016/S1473-3099(16)00078-5 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
