

Phase separation of polyubiquitinated proteins in UBQLN2 condensates controls substrate fate
- PMID: 39121161
- PMCID: PMC11331126
- DOI: 10.1073/pnas.2405964121
Phase separation of polyubiquitinated proteins in UBQLN2 condensates controls substrate fate
Abstract
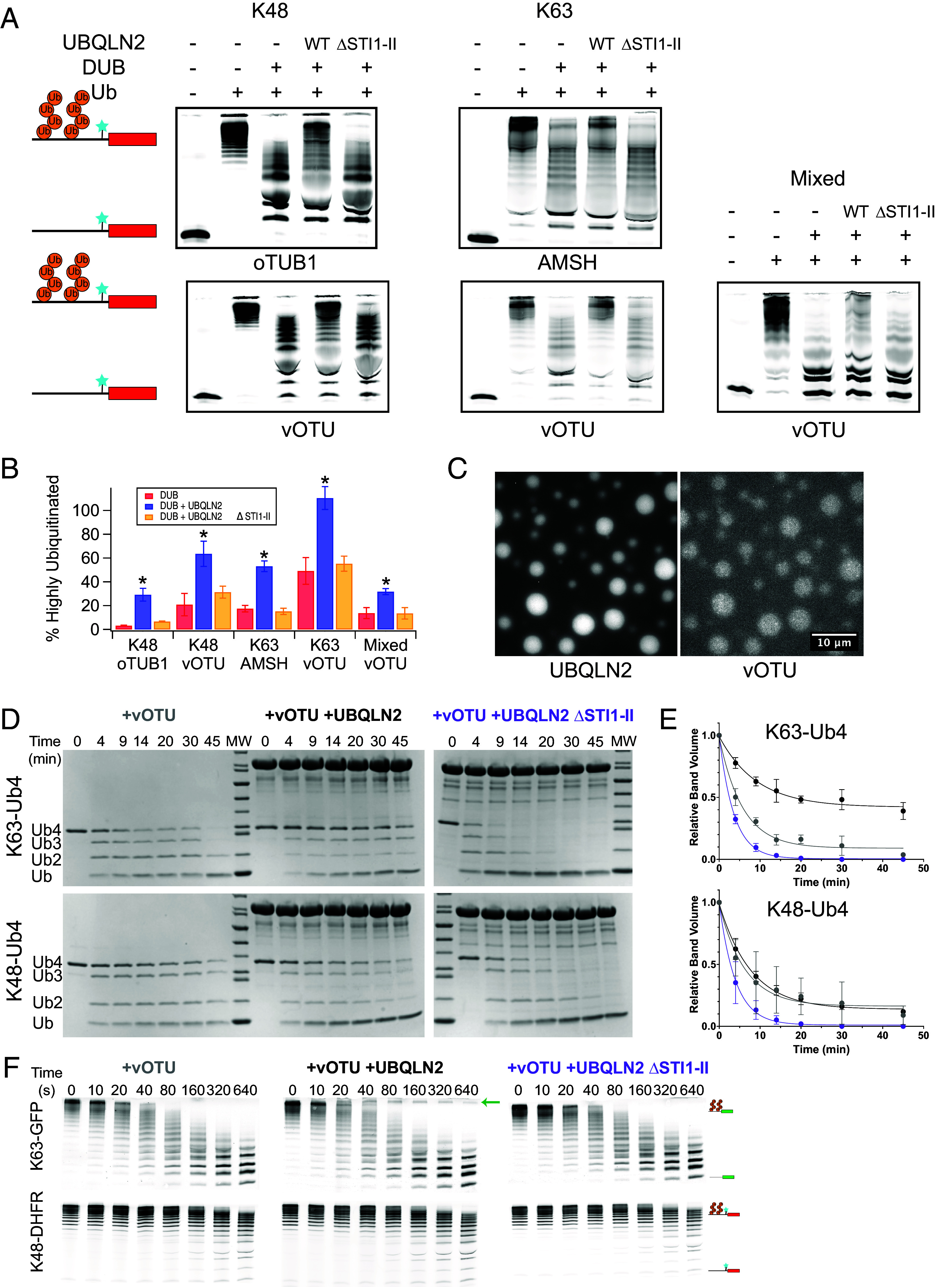
Ubiquitination is one of the most common posttranslational modifications in eukaryotic cells. Depending on the architecture of polyubiquitin chains, substrate proteins can meet different cellular fates, but our understanding of how chain linkage controls protein fate remains limited. UBL-UBA shuttle proteins, such as UBQLN2, bind to ubiquitinated proteins and to the proteasome or other protein quality control machinery elements and play a role in substrate fate determination. Under physiological conditions, UBQLN2 forms biomolecular condensates through phase separation, a physicochemical phenomenon in which multivalent interactions drive the formation of a macromolecule-rich dense phase. Ubiquitin and polyubiquitin chains modulate UBQLN2's phase separation in a linkage-dependent manner, suggesting a possible link to substrate fate determination, but polyubiquitinated substrates have not been examined directly. Using sedimentation assays and microscopy we show that polyubiquitinated substrates induce UBQLN2 phase separation and incorporate into the resulting condensates. This substrate effect is strongest with K63-linked substrates, intermediate with mixed-linkage substrates, and weakest with K48-linked substrates. Proteasomes can be recruited to these condensates, but proteasome activity toward K63-linked and mixed linkage substrates is inhibited in condensates. Substrates are also protected from deubiquitinases by UBQLN2-induced phase separation. Our results suggest that phase separation could regulate the fate of ubiquitinated substrates in a chain-linkage-dependent manner, thus serving as an interpreter of the ubiquitin code.
Keywords: Condensates; phase separation; proteasome; protein quality control; ubiquitin.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures




Update of
-
Phase separation of polyubiquitinated proteins in UBQLN2 condensates controls substrate fate.bioRxiv [Preprint]. 2024 Jun 20:2024.03.15.585243. doi: 10.1101/2024.03.15.585243. bioRxiv. 2024. Update in: Proc Natl Acad Sci U S A. 2024 Aug 13;121(33):e2405964121. doi: 10.1073/pnas.2405964121. PMID: 38559018 Free PMC article. Updated. Preprint.
Similar articles
-
Phase separation of polyubiquitinated proteins in UBQLN2 condensates controls substrate fate.bioRxiv [Preprint]. 2024 Jun 20:2024.03.15.585243. doi: 10.1101/2024.03.15.585243. bioRxiv. 2024. Update in: Proc Natl Acad Sci U S A. 2024 Aug 13;121(33):e2405964121. doi: 10.1073/pnas.2405964121. PMID: 38559018 Free PMC article. Updated. Preprint.
-
Ubiquitin Modulates Liquid-Liquid Phase Separation of UBQLN2 via Disruption of Multivalent Interactions.Mol Cell. 2018 Mar 15;69(6):965-978.e6. doi: 10.1016/j.molcel.2018.02.004. Epub 2018 Mar 8. Mol Cell. 2018. PMID: 29526694 Free PMC article.
-
Polyubiquitin ligand-induced phase transitions are optimized by spacing between ubiquitin units.Proc Natl Acad Sci U S A. 2023 Oct 17;120(42):e2306638120. doi: 10.1073/pnas.2306638120. Epub 2023 Oct 12. Proc Natl Acad Sci U S A. 2023. PMID: 37824531 Free PMC article.
-
The recognition of ubiquitinated proteins by the proteasome.Cell Mol Life Sci. 2016 Sep;73(18):3497-506. doi: 10.1007/s00018-016-2255-5. Epub 2016 May 2. Cell Mol Life Sci. 2016. PMID: 27137187 Free PMC article. Review.
-
Ubiquitin-Modulated Phase Separation of Shuttle Proteins: Does Condensate Formation Promote Protein Degradation?Bioessays. 2020 Nov;42(11):e2000036. doi: 10.1002/bies.202000036. Epub 2020 Sep 3. Bioessays. 2020. PMID: 32881044 Free PMC article. Review.
Cited by
-
UbiREAD deciphers proteasomal degradation code of homotypic and branched K48 and K63 ubiquitin chains.Mol Cell. 2025 Apr 3;85(7):1467-1476.e6. doi: 10.1016/j.molcel.2025.02.021. Epub 2025 Mar 24. Mol Cell. 2025. PMID: 40132582 Free PMC article.
-
STI1 domain dynamically engages transient helices in disordered regions to drive self-association and phase separation of yeast ubiquilin Dsk2.bioRxiv [Preprint]. 2025 May 13:2025.03.14.643327. doi: 10.1101/2025.03.14.643327. bioRxiv. 2025. PMID: 40161686 Free PMC article. Preprint.
-
Chronic Oxidative Stress and Stress Granule Formation in UBQLN2 ALS Neurons: Insights into Neuronal Degeneration and Potential Therapeutic Targets.Int J Mol Sci. 2024 Dec 15;25(24):13448. doi: 10.3390/ijms252413448. Int J Mol Sci. 2024. PMID: 39769213 Free PMC article.
-
Protein quality control machinery: regulators of condensate architecture and functionality.Trends Biochem Sci. 2025 Feb;50(2):106-120. doi: 10.1016/j.tibs.2024.12.003. Epub 2025 Jan 3. Trends Biochem Sci. 2025. PMID: 39755440 Review.
-
Endogenous retrovirus-like proteins recruit UBQLN2 to stress granules and shape their functional biology.Sci Adv. 2025 Jul 18;11(29):eadu6354. doi: 10.1126/sciadv.adu6354. Epub 2025 Jul 18. Sci Adv. 2025. PMID: 40680123 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
