

Cyclin F-EXO1 axis controls cell cycle-dependent execution of double-strand break repair
- PMID: 39121215
- PMCID: PMC11313846
- DOI: 10.1126/sciadv.ado0636
Cyclin F-EXO1 axis controls cell cycle-dependent execution of double-strand break repair
Abstract
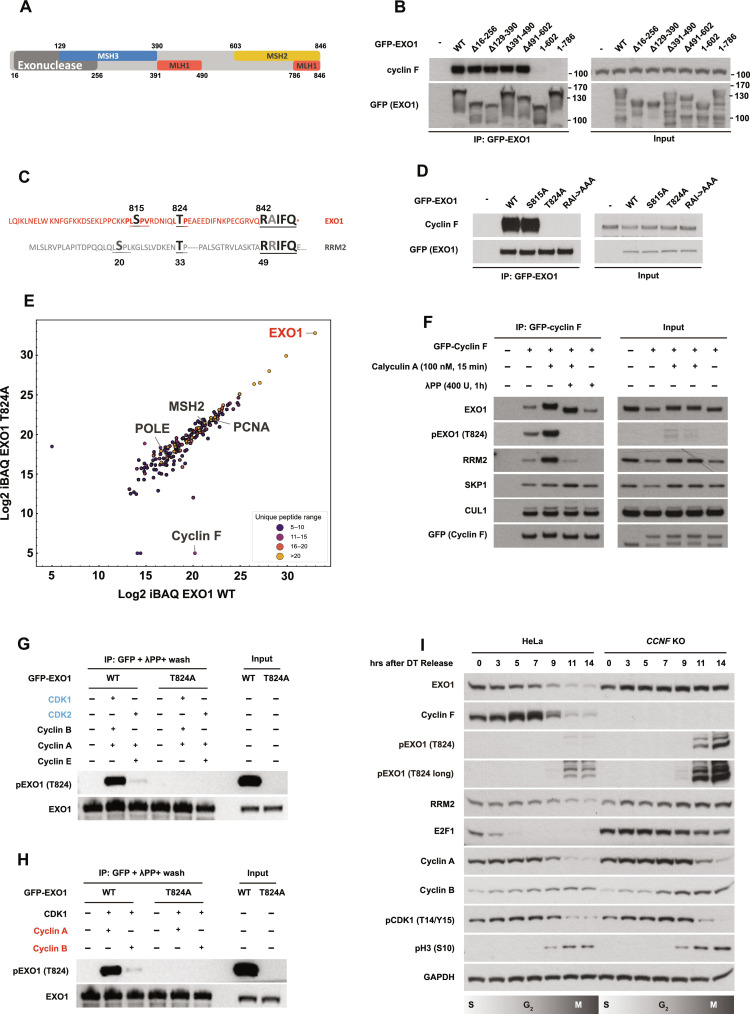
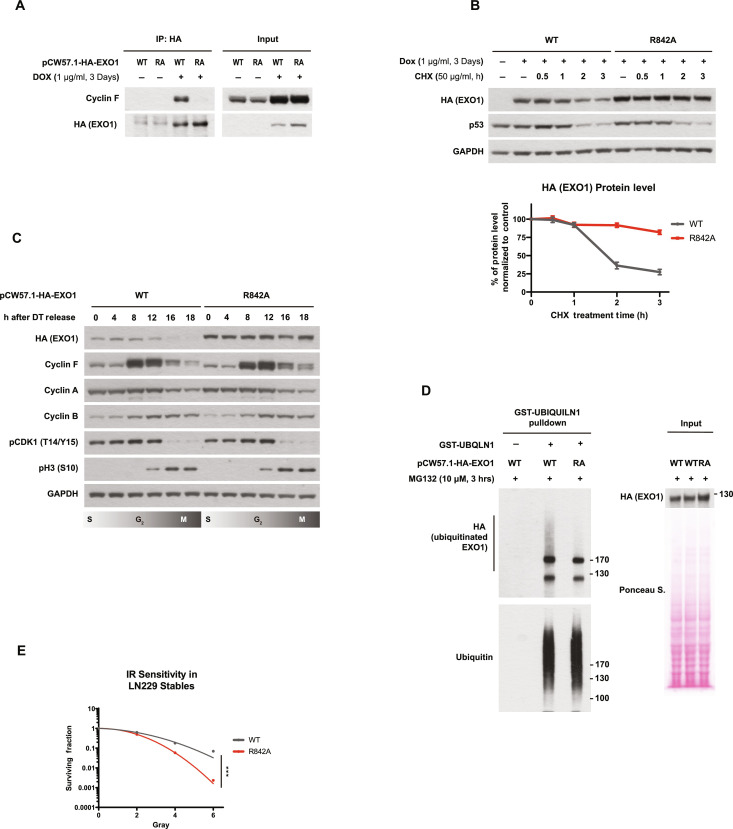
Ubiquitination is a crucial posttranslational modification required for the proper repair of DNA double-strand breaks (DSBs) induced by ionizing radiation (IR). DSBs are mainly repaired through homologous recombination (HR) when template DNA is present and nonhomologous end joining (NHEJ) in its absence. In addition, microhomology-mediated end joining (MMEJ) and single-strand annealing (SSA) provide backup DSBs repair pathways. However, the mechanisms controlling their use remain poorly understood. By using a high-resolution CRISPR screen of the ubiquitin system after IR, we systematically uncover genes required for cell survival and elucidate a critical role of the E3 ubiquitin ligase SCFcyclin F in cell cycle-dependent DSB repair. We show that SCFcyclin F-mediated EXO1 degradation prevents DNA end resection in mitosis, allowing MMEJ to take place. Moreover, we identify a conserved cyclin F recognition motif, distinct from the one used by other cyclins, with broad implications in cyclin specificity for cell cycle control.
Figures





References
-
- Schwertman P., Bekker-Jensen S., Mailand N., Regulation of DNA double-strand break repair by ubiquitin and ubiquitin-like modifiers. Nat. Rev. Mol. Cell Biol. 17, 379–394 (2016). - PubMed
-
- Chapman J. R., Taylor M. R., Boulton S. J., Playing the end game: DNA double-strand break repair pathway choice. Mol. Cell 47, 497–510 (2012). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
