

Integrated single-cell RNA-seq analysis reveals mitochondrial calcium signaling as a modulator of endothelial-to-mesenchymal transition
- PMID: 39121218
- PMCID: PMC11313856
- DOI: 10.1126/sciadv.adp6182
Integrated single-cell RNA-seq analysis reveals mitochondrial calcium signaling as a modulator of endothelial-to-mesenchymal transition
Abstract
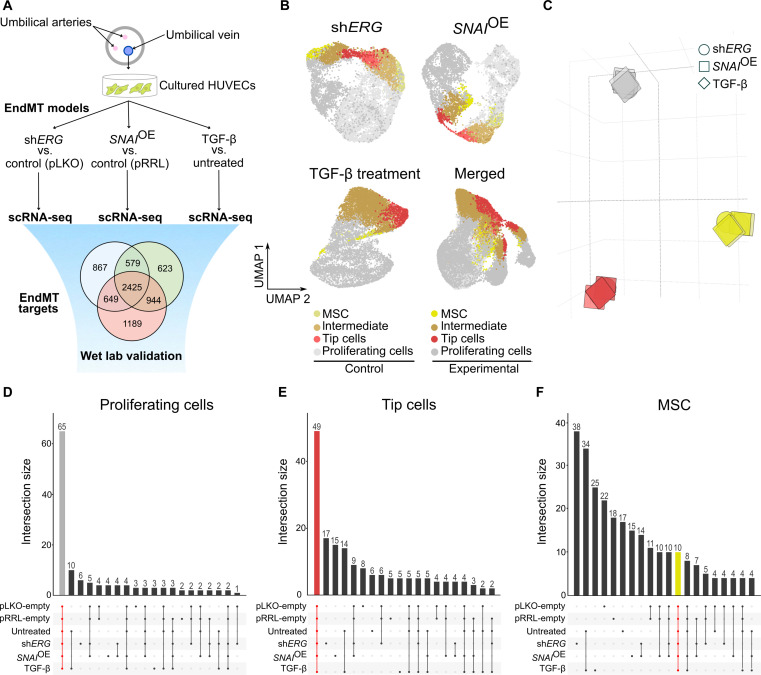
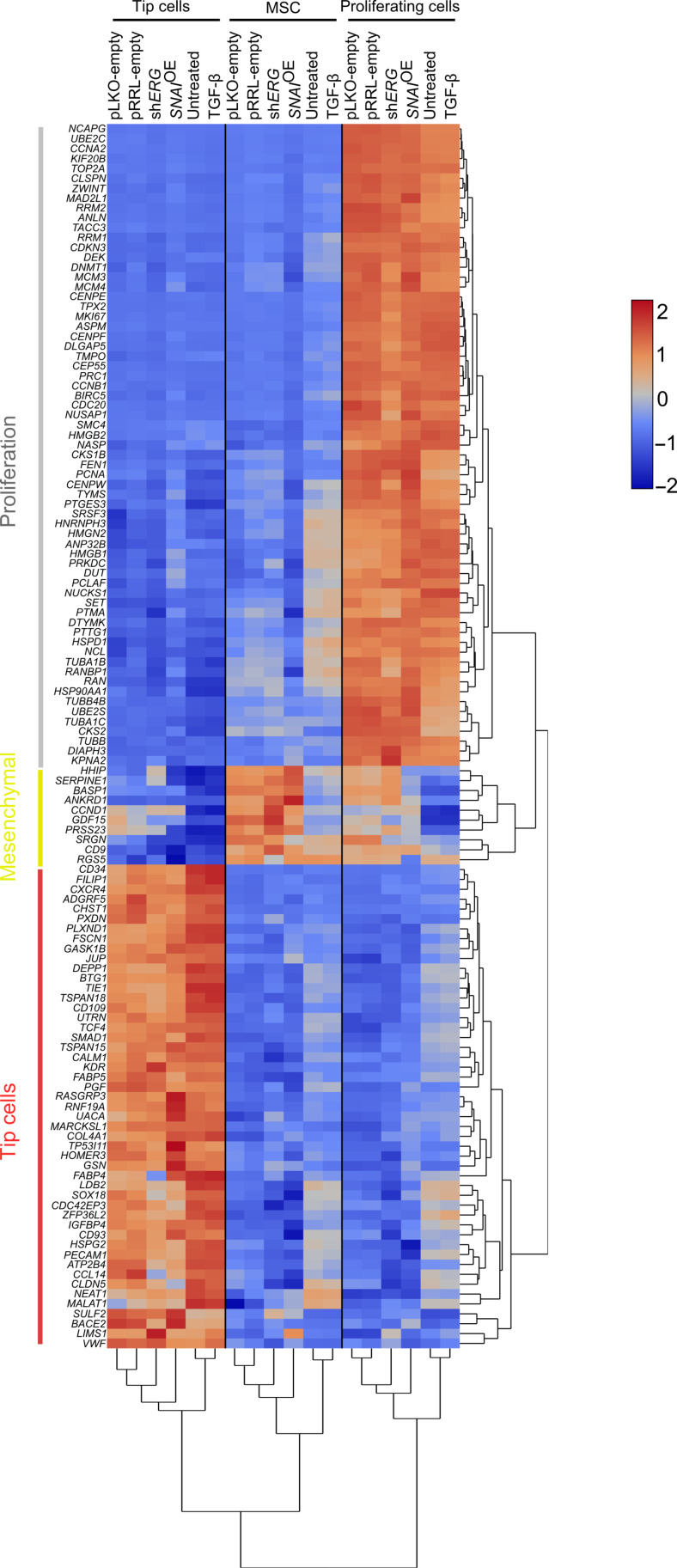
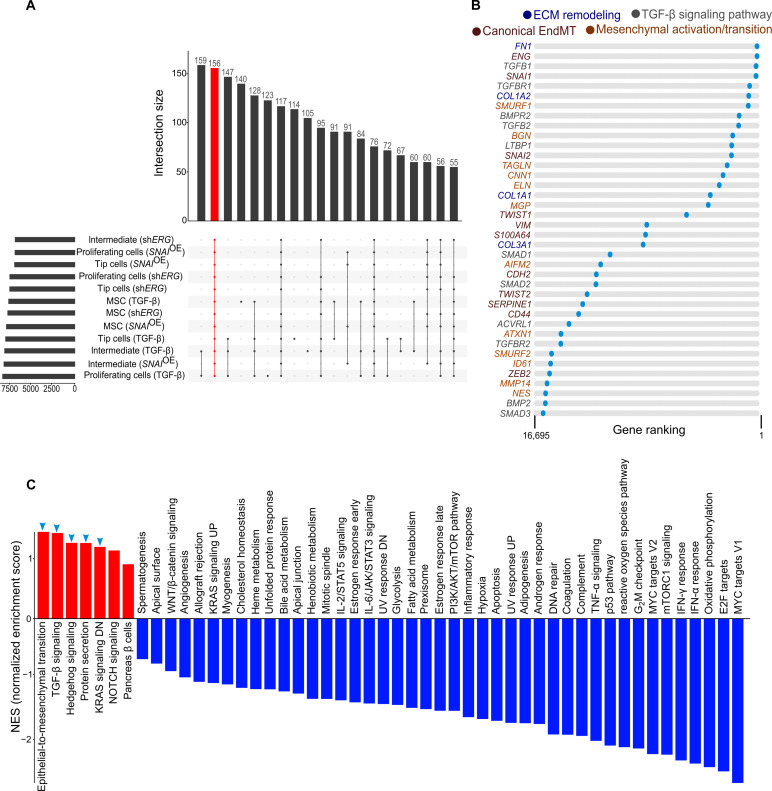
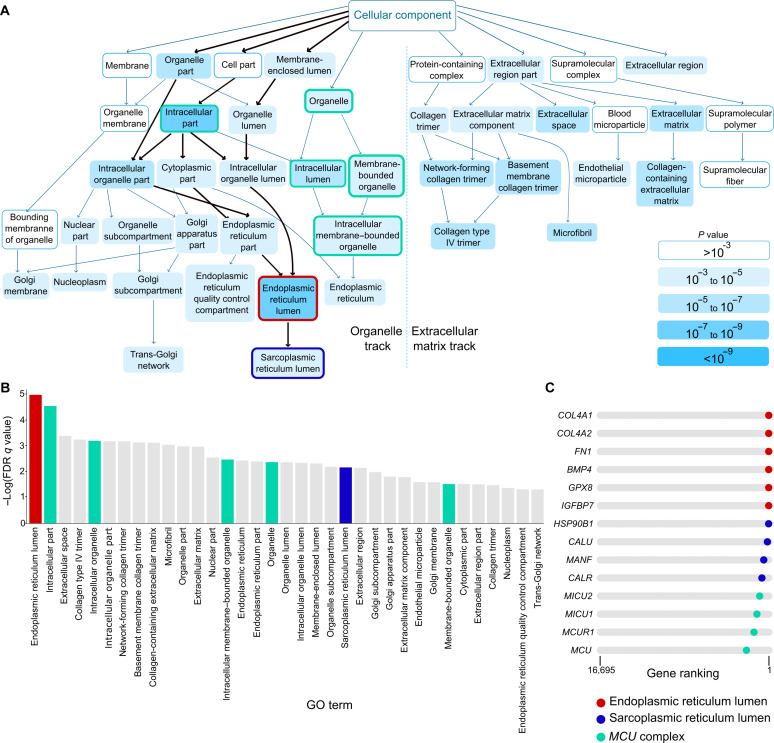
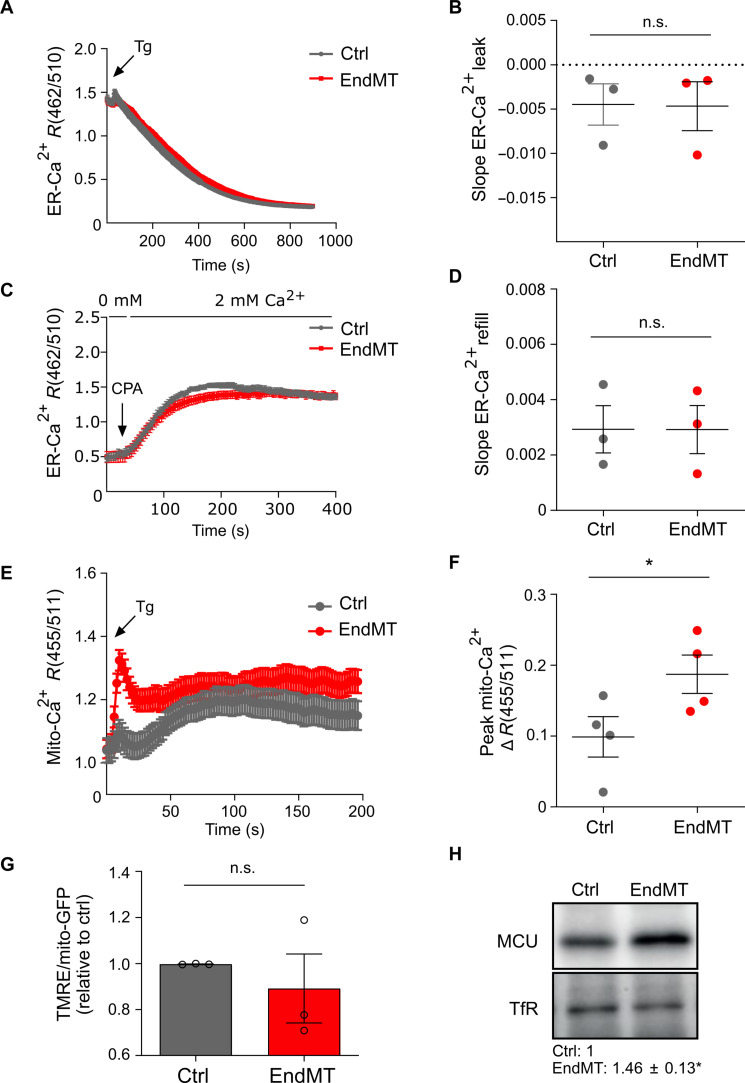
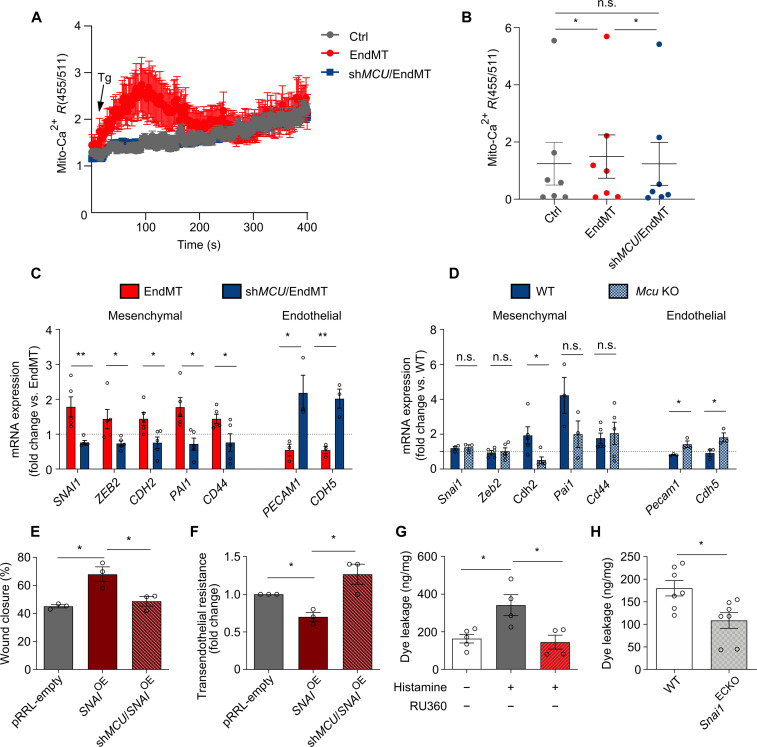
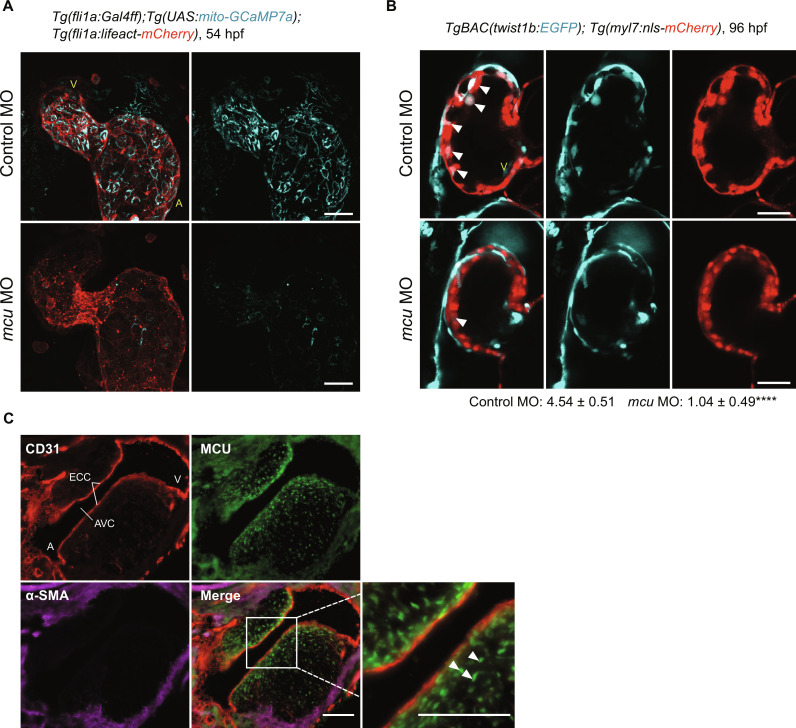
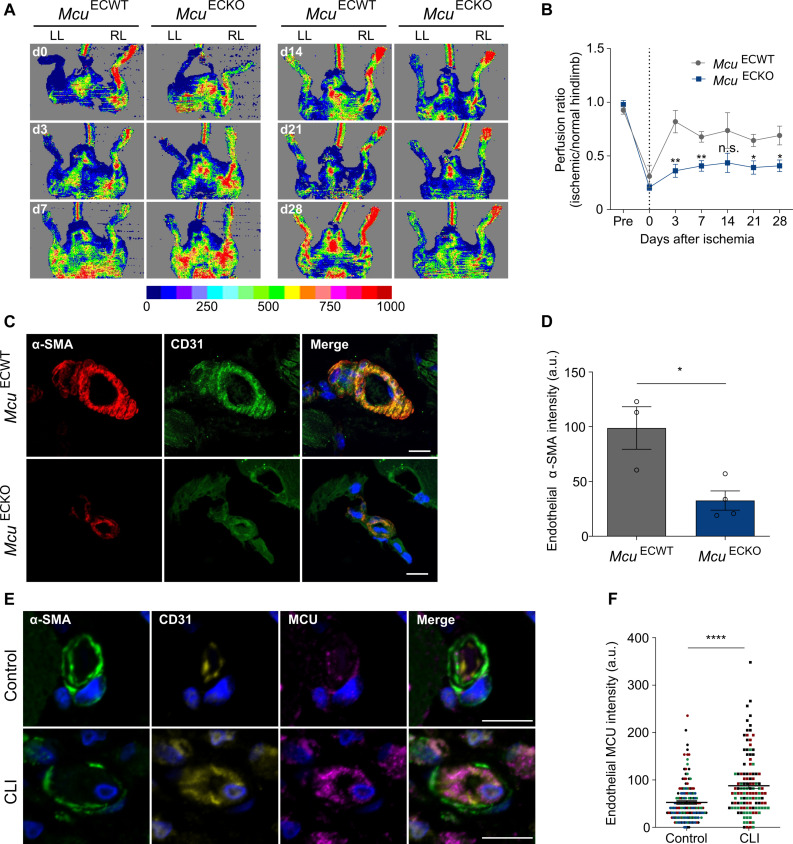
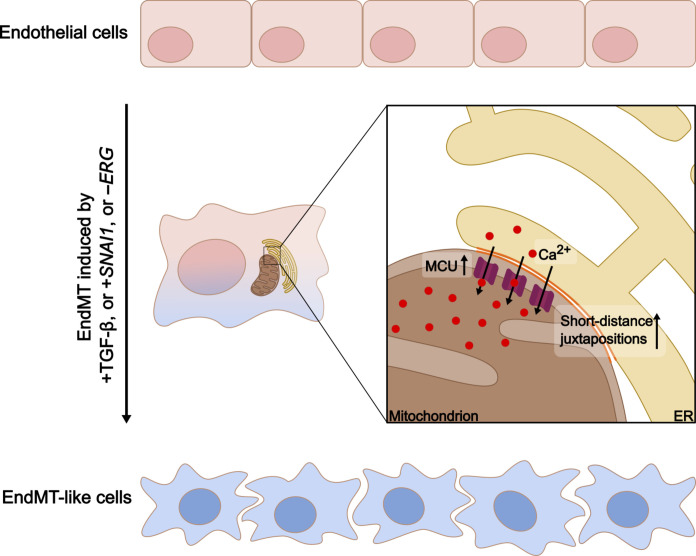
Endothelial cells (ECs) are highly plastic, capable of differentiating into various cell types. Endothelial-to-mesenchymal transition (EndMT) is crucial during embryonic development and contributes substantially to vascular dysfunction in many cardiovascular diseases (CVDs). While targeting EndMT holds therapeutic promise, understanding its mechanisms and modulating its pathways remain challenging. Using single-cell RNA sequencing on three in vitro EndMT models, we identified conserved gene signatures. We validated original regulators in vitro and in vivo during embryonic heart development and peripheral artery disease. EndMT induction led to global expression changes in all EC subtypes rather than in mesenchymal clusters. We identified mitochondrial calcium uptake as a key driver of EndMT; inhibiting mitochondrial calcium uniporter (MCU) prevented EndMT in vitro, and conditional Mcu deletion in ECs blocked mesenchymal activation in a hind limb ischemia model. Tissues from patients with critical limb ischemia with EndMT features exhibited significantly elevated endothelial MCU. These findings highlight MCU as a regulator of EndMT and a potential therapeutic target.
Figures
References
-
- Sanchez-Duffhues G., Orlova V., Ten Dijke P., In brief: Endothelial-to-mesenchymal transition. J. Pathol. 238, 378–380 (2016). - PubMed
-
- Anderberg C., Cunha S. I., Zhai Z., Cortez E., Pardali E., Johnson J. R., Franco M., Paez-Ribes M., Cordiner R., Fuxe J., Johansson B. R., Goumans M. J., Casanovas O., ten Dijke P., Arthur H. M., Pietras K., Deficiency for endoglin in tumor vsculature weakens the endothelial barrier to metastatic dissemination. J. Exp. Med. 210, 563–579 (2013). - PMC - PubMed
-
- Lovisa S., Fletcher-Sananikone E., Sugimoto H., Hensel J., Lahiri S., Hertig A., Taduri G., Lawson E., Dewar R., Revuelta I., Kato N., Wu C. J., Bassett R. L. Jr., Putluri N., Zeisberg M., Zeisberg E. M., LeBleu V. S., Kalluri R., Endothelial-to-mesenchymal transition compromises vascular integrity to induce Myc-mediated metabolic reprogramming in kidney fibrosis. Sci. Signal. 13, eaaz2597 (2020). - PMC - PubMed
-
- Arciniegas E., Sutton A. B., Allen T. D., Schor A. M., Transforming growth factor beta 1 promotes the differentiation of endothelial cells into smooth muscle-like cells in vitro. J. Cell Sci. 103, 521–529 (1992). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
