

Guidelines for minimal information on cellular senescence experimentation in vivo
- PMID: 39121846
- PMCID: PMC11790242
- DOI: 10.1016/j.cell.2024.05.059
Guidelines for minimal information on cellular senescence experimentation in vivo
Abstract
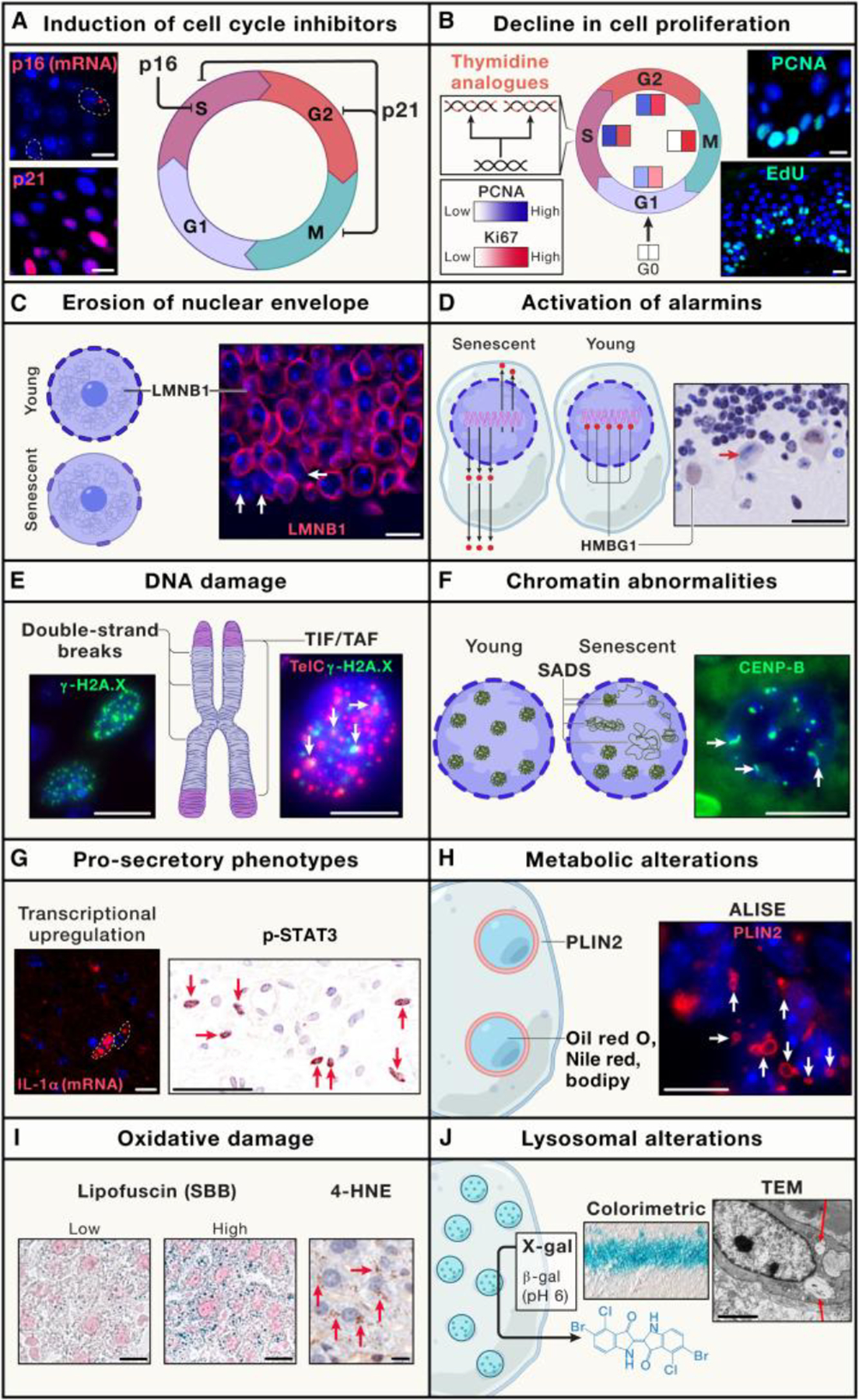
Cellular senescence is a cell fate triggered in response to stress and is characterized by stable cell-cycle arrest and a hypersecretory state. It has diverse biological roles, ranging from tissue repair to chronic disease. The development of new tools to study senescence in vivo has paved the way for uncovering its physiological and pathological roles and testing senescent cells as a therapeutic target. However, the lack of specific and broadly applicable markers makes it difficult to identify and characterize senescent cells in tissues and living organisms. To address this, we provide practical guidelines called "minimum information for cellular senescence experimentation in vivo" (MICSE). It presents an overview of senescence markers in rodent tissues, transgenic models, non-mammalian systems, human tissues, and tumors and their use in the identification and specification of senescent cells. These guidelines provide a uniform, state-of-the-art, and accessible toolset to improve our understanding of cellular senescence in vivo.
Keywords: aging; humans; in vivo; mouse; senescence; senotherapy.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests D.J.B. has potential financial interests related to this study. He is a co-inventor of patents held by the Mayo Clinic, patent applications licensed to or filed by Unity Biotechnology, and a Unity Biotechnology shareholder. Research in the Baker laboratory has been reviewed by the Mayo Clinic Conflict of Interest Review Board and is being conducted in compliance with Mayo Clinic Conflict of Interest policies. J. Gil has acted as a consultant for Unity Biotechnology, Geras Bio, Myricx Pharma Ltd., and Merck KGaA; owns equity in Geras Bio and share options in Myricx Pharma Ltd.; and is a named inventor in MRC and Imperial College patents related to senolytic therapies. J. Gil currently receives funding from Pfizer. Unity Biotechnology funded research on senolytics in J. Gil’s laboratory in the past. SenTraGor and GLF16 senescence detection compounds are under patent applications: EP3475287B1, and 20240100309 (Greek patent application) along with GB2406749.8 (UK patent application), respectively. J.M.S. is a co-inventor on patents held by Brown University on methods to inhibit retrotransposon activation in age-related diseases. He is the scientific co-founder of Transposon Therapeutics, chair of their scientific advisory board, and a consultant and holds stock options. He is also a consultant and holds equity in Atropos Therapeutics. Research in the Sedivy laboratory has been reviewed by the Brown University Conflict of Interest Review Board and is being conducted in compliance with Brown University Conflict of Interest policies. F.d.d.F. is an inventor on the patent applications PCT/EP2013/059753 and PCT/EP2016/068162. M.D. is co-inventor on patents held by the Buck Institute for Research on Aging. He is the scientific co-founder of Cleara Biotech and consultant for Oisin Biotechnologies. M.D.’s laboratory currently receives research funding from Ono Pharmaceuticals. J. Grillari is co-inventor on patents held by BOKU and is a co-founder and scientific advisor to TAmiRNA and Rockfish Bio.
Figures



References
-
- Thery C, Witwer KW, Aikawa E, Alcaraz MJ, Anderson JD, Andriantsitohaina R, Antoniou A, Arab T, Archer F, Atkin-Smith GK, et al. (2018). Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles 7, 1535750. 10.1080/20013078.2018.1535750. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U54 AG076041/AG/NIA NIH HHS/United States
- R01 AG068076/AG/NIA NIH HHS/United States
- R01 AG053229/AG/NIA NIH HHS/United States
- R37 AG013925/AG/NIA NIH HHS/United States
- R01 AG071861/AG/NIA NIH HHS/United States
- U54 AG079779/AG/NIA NIH HHS/United States
- U19 AG056278/AG/NIA NIH HHS/United States
- R01 AG063543/AG/NIA NIH HHS/United States
- R01 AG068048/AG/NIA NIH HHS/United States
- U54 AG079758/AG/NIA NIH HHS/United States
- UG3 CA268103/CA/NCI NIH HHS/United States
- R33 AG061456/AG/NIA NIH HHS/United States
- P01 AG043376/AG/NIA NIH HHS/United States
- P01 AG031862/AG/NIA NIH HHS/United States
- U54 AG079754/AG/NIA NIH HHS/United States
- UH3 CA268103/CA/NCI NIH HHS/United States
- P01 AG062413/AG/NIA NIH HHS/United States
- R01 AG082708/AG/NIA NIH HHS/United States
- R01 AG068182/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
