

Enhancing homology-directed repair efficiency with HDR-boosting modular ssDNA donor
- PMID: 39122671
- PMCID: PMC11315919
- DOI: 10.1038/s41467-024-50788-x
Enhancing homology-directed repair efficiency with HDR-boosting modular ssDNA donor
Abstract
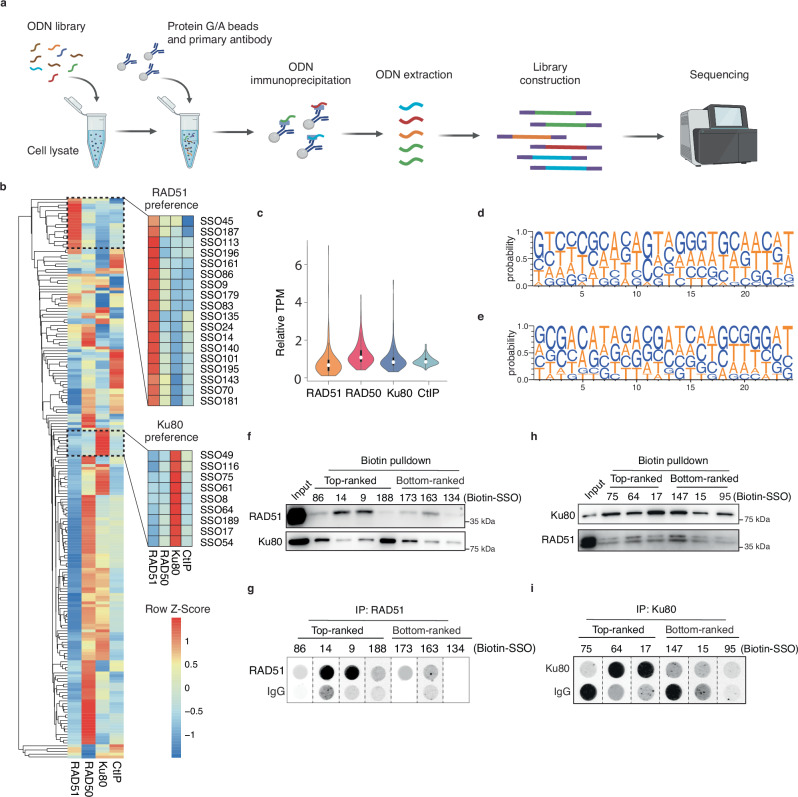
Despite the potential of small molecules and recombinant proteins to enhance the efficiency of homology-directed repair (HDR), single-stranded DNA (ssDNA) donors, as currently designed and chemically modified, remain suboptimal for precise gene editing. Here, we screen the biased ssDNA binding sequences of DNA repair-related proteins and engineer RAD51-preferred sequences into HDR-boosting modules for ssDNA donors. Donors with these modules exhibit an augmented affinity for RAD51, thereby enhancing HDR efficiency across various genomic loci and cell types when cooperated with Cas9, nCas9, and Cas12a. By combining with an inhibitor of non-homologous end joining (NHEJ) or the HDRobust strategy, these modular ssDNA donors achieve up to 90.03% (median 74.81%) HDR efficiency. The HDR-boosting modules targeting an endogenous protein enable a chemical modification-free strategy to improve the efficacy of ssDNA donors for precise gene editing.
© 2024. The Author(s).
Conflict of interest statement
D.L., Y.J., P.Z., and H.C. have submitted two patent applications to the China National Intellectual Property Administration pertaining to the HDR-boosting modules and the combination of HDR-boosting modular ssDNA donor and M3814 inhibitor aspects of this work (application number 2023108476157 and 2024107260446). The remaining authors declare no competing interests.
Figures





References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
