

High-throughput identification of functional regulatory SNPs in systemic lupus erythematosus
- PMID: 39122710
- PMCID: PMC11315931
- DOI: 10.1038/s41467-024-50710-5
High-throughput identification of functional regulatory SNPs in systemic lupus erythematosus
Abstract
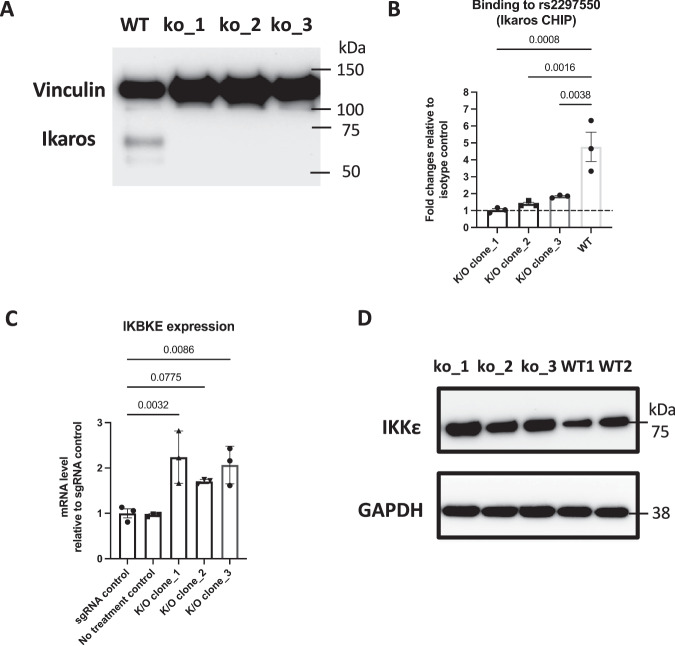
Genome-wide association studies implicate multiple loci in risk for systemic lupus erythematosus (SLE), but few contain exonic variants, rendering systematic identification of non-coding variants essential to decoding SLE genetics. We utilized SNP-seq and bioinformatic enrichment to interrogate 2180 single-nucleotide polymorphisms (SNPs) from 87 SLE risk loci for potential binding of transcription factors and related proteins from B cells. 52 SNPs that passed initial screening were tested by electrophoretic mobility shift and luciferase reporter assays. To validate the approach, we studied rs2297550 in detail, finding that the risk allele enhanced binding to the transcription factor Ikaros (encoded by IKZF1), thereby modulating expression of IKBKE. Correspondingly, primary cells from genotyped healthy donors bearing the risk allele expressed higher levels of the interferon / NF-κB regulator IKKε. Together, these findings define a set of likely functional non-coding lupus risk variants and identify a regulatory pathway involving rs2297550, Ikaros, and IKKε implicated by human genetics in risk for SLE.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
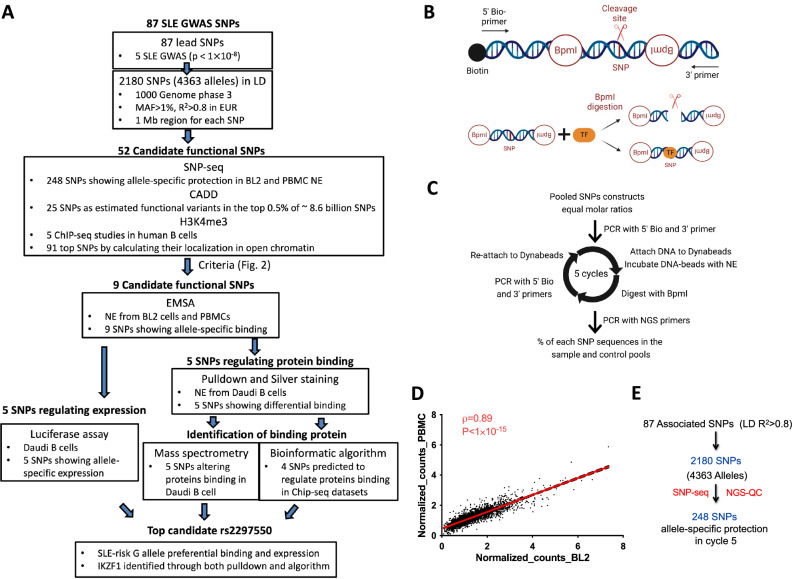
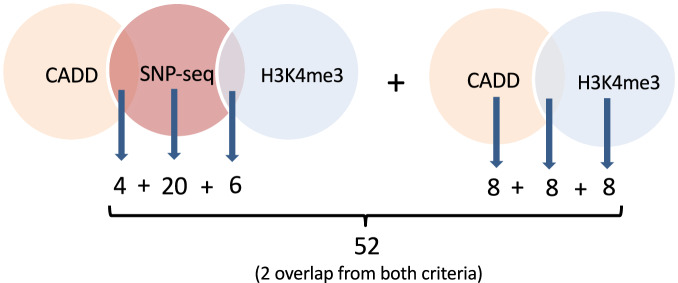
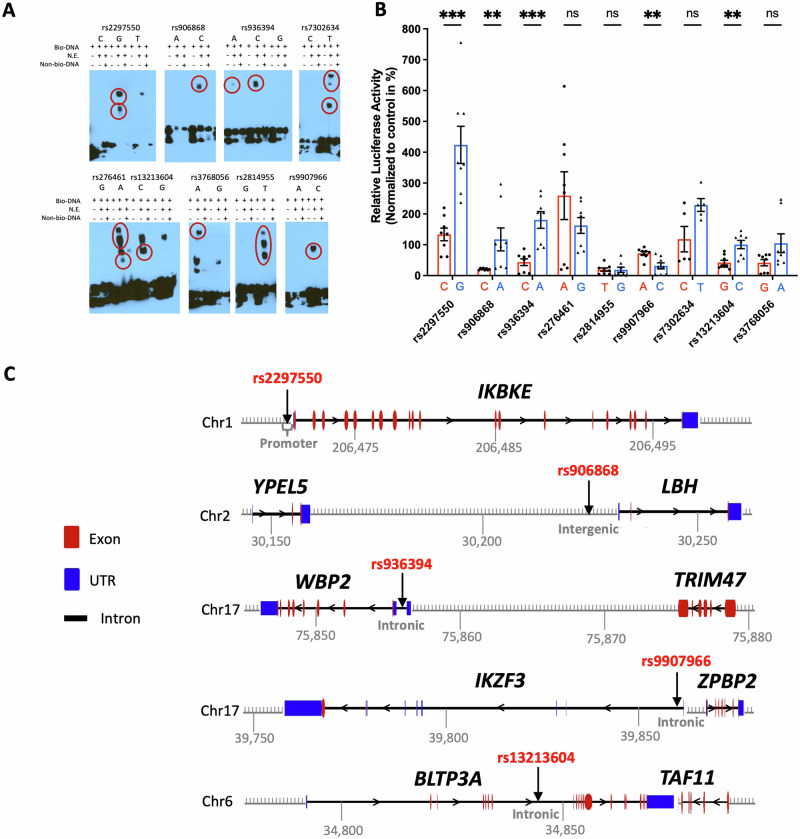
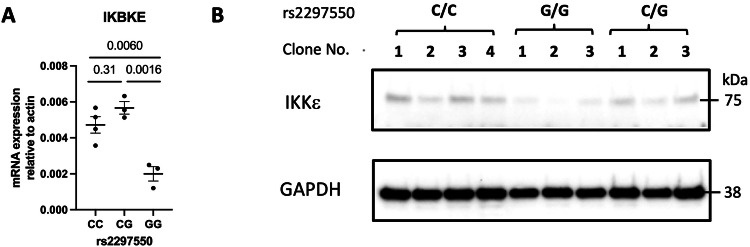



Update of
-
High-throughput identification of functional regulatory SNPs in systemic lupus erythematosus.bioRxiv [Preprint]. 2024 Apr 12:2023.08.16.553538. doi: 10.1101/2023.08.16.553538. bioRxiv. 2024. Update in: Nat Commun. 2024 Aug 9;15(1):6804. doi: 10.1038/s41467-024-50710-5. PMID: 37645953 Free PMC article. Updated. Preprint.
References
-
- Vinuesa, C. G., Shen, N. & Ware, T. Genetics of SLE: mechanistic insights from monogenic disease and disease-associated variants. Nat. Rev. Nephrol. 19, 558–572 (2023). - PubMed
MeSH terms
Substances
Grants and funding
- 2R01AR065538/U.S. Department of Health & Human Services | NIH | National Institute of Arthritis and Musculoskeletal and Skin Diseases (NIAMS)
- U01 AI130830/AI/NIAID NIH HHS/United States
- R01 AR073228/AR/NIAMS NIH HHS/United States
- R01 NS099068/NS/NINDS NIH HHS/United States
- P30 AR072577/AR/NIAMS NIH HHS/United States
- R01 AR073201/AR/NIAMS NIH HHS/United States
- P30 AR070253/AR/NIAMS NIH HHS/United States
- R01 AR065538/AR/NIAMS NIH HHS/United States
- R01 HG010730/HG/NHGRI NIH HHS/United States
- T32 AR007611/AR/NIAMS NIH HHS/United States
- P30AR070253/U.S. Department of Health & Human Services | NIH | National Institute of Arthritis and Musculoskeletal and Skin Diseases (NIAMS)
- R01 AI024717/AI/NIAID NIH HHS/United States
- P30 AR070549/AR/NIAMS NIH HHS/United States
- R01 AR075906/AR/NIAMS NIH HHS/United States
- P30 AR069625/AR/NIAMS NIH HHS/United States
- R01 AR077607/AR/NIAMS NIH HHS/United States
- R01AR073201/U.S. Department of Health & Human Services | NIH | National Institute of Arthritis and Musculoskeletal and Skin Diseases (NIAMS)
- R01AR075906/U.S. Department of Health & Human Services | NIH | National Institute of Arthritis and Musculoskeletal and Skin Diseases (NIAMS)
LinkOut - more resources
Full Text Sources
Medical
