

The Foxo1-YAP-Notch1 axis reprograms STING-mediated innate immunity in NASH progression
- PMID: 39122845
- PMCID: PMC11372114
- DOI: 10.1038/s12276-024-01280-5
The Foxo1-YAP-Notch1 axis reprograms STING-mediated innate immunity in NASH progression
Abstract
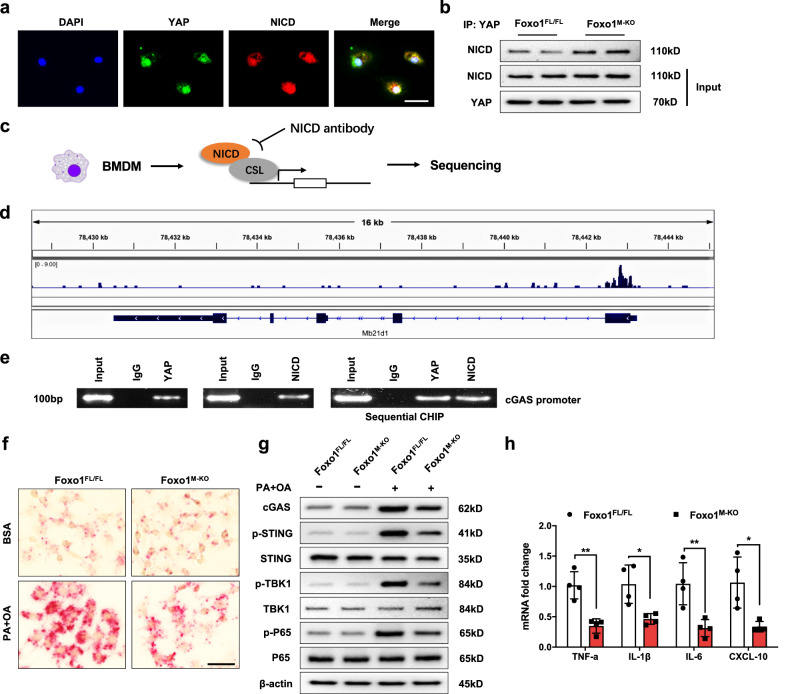
Innate immune activation is critical for initiating hepatic inflammation during nonalcoholic steatohepatitis (NASH) progression. However, the mechanisms by which immunoregulatory molecules recognize lipogenic, fibrotic, and inflammatory signals remain unclear. Here, we show that high-fat diet (HFD)-induced oxidative stress activates Foxo1, YAP, and Notch1 signaling in hepatic macrophages. Macrophage Foxo1 deficiency (Foxo1M-KO) ameliorated hepatic inflammation, steatosis, and fibrosis, with reduced STING, TBK1, and NF-κB activation in HFD-challenged livers. However, Foxo1 and YAP double knockout (Foxo1/YAPM-DKO) or Foxo1 and Notch1 double knockout (Foxo1/Notch1M-DKO) promoted STING function and exacerbated HFD-induced liver injury. Interestingly, Foxo1M-KO strongly reduced TGF-β1 release from palmitic acid (PA)- and oleic acid (OA)-stimulated Kupffer cells and decreased Col1α1, CCL2, and Timp1 expression but increased MMP1 expression in primary hepatic stellate cells (HSCs) after coculture with Kupffer cells. Notably, PA and OA challenge in Kupffer cells augmented LIMD1 and LATS1 colocalization and interaction, which induced YAP nuclear translocation. Foxo1M-KO activated PGC-1α and increased nuclear YAP activity, modulating mitochondrial biogenesis. Using chromatin immunoprecipitation (ChIP) coupled with massively parallel sequencing (ChIP-Seq) and in situ RNA hybridization, we found that NICD colocalizes with YAP and targets Mb21d1 (cGAS), while YAP functions as a novel coactivator of the NICD, which is crucial for reprogramming STING function in NASH progression. These findings highlight the importance of the macrophage Foxo1-YAP-Notch1 axis as a key molecular regulator that controls lipid metabolism, inflammation, and innate immunity in NASH.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures







References
MeSH terms
Substances
Grants and funding
- R01AI139552/Foundation for the National Institutes of Health (Foundation for the National Institutes of Health, Inc.)
- R21 AI146742/AI/NIAID NIH HHS/United States
- R21 AI112722/AI/NIAID NIH HHS/United States
- R21AI112722/Foundation for the National Institutes of Health (Foundation for the National Institutes of Health, Inc.)
- R01 AI139552/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
