

Mitochondrial permeability transition dictates mitochondrial maturation upon switch in cellular identity of hematopoietic precursors
- PMID: 39122870
- PMCID: PMC11316084
- DOI: 10.1038/s42003-024-06671-y
Mitochondrial permeability transition dictates mitochondrial maturation upon switch in cellular identity of hematopoietic precursors
Abstract
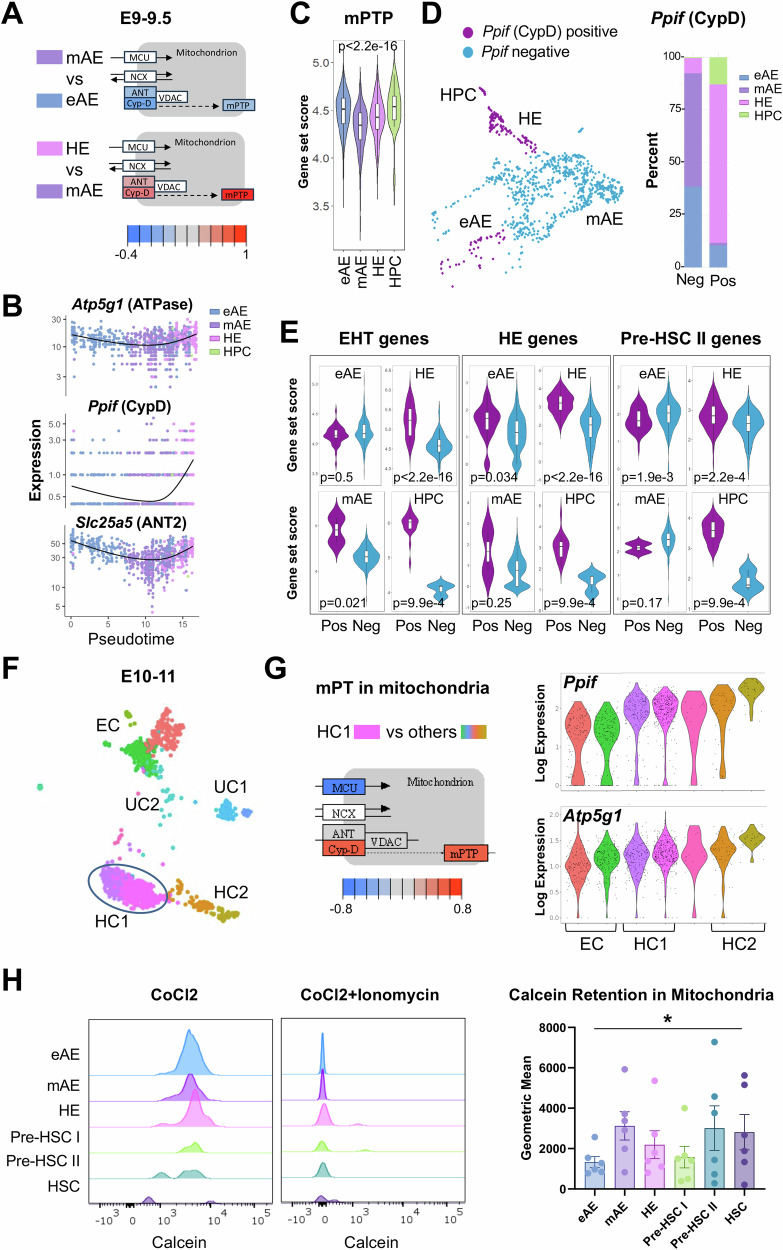
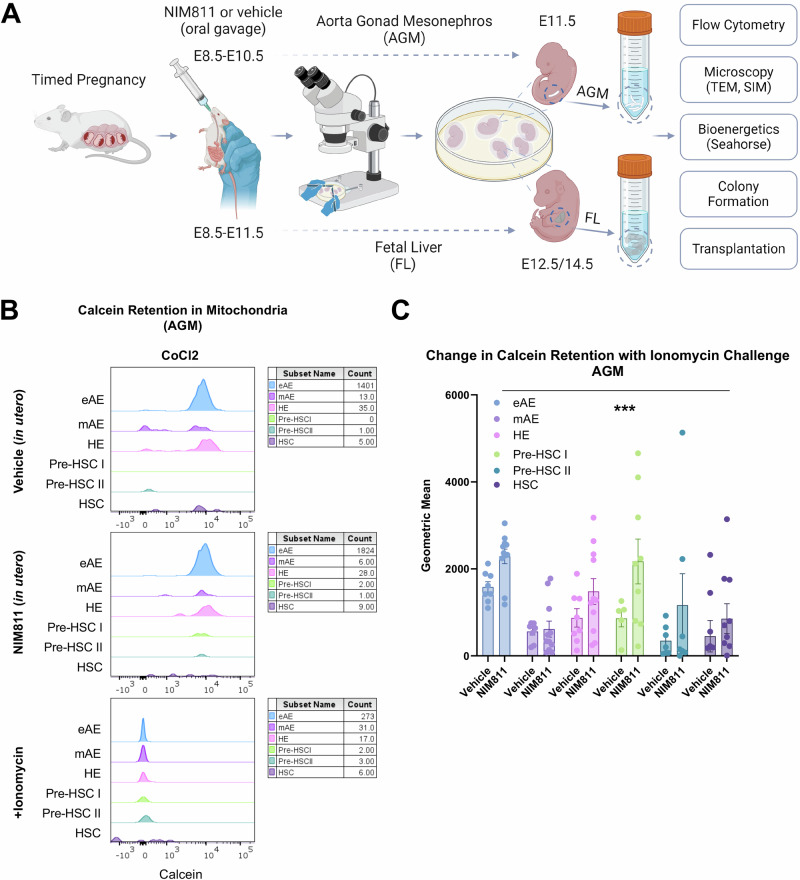
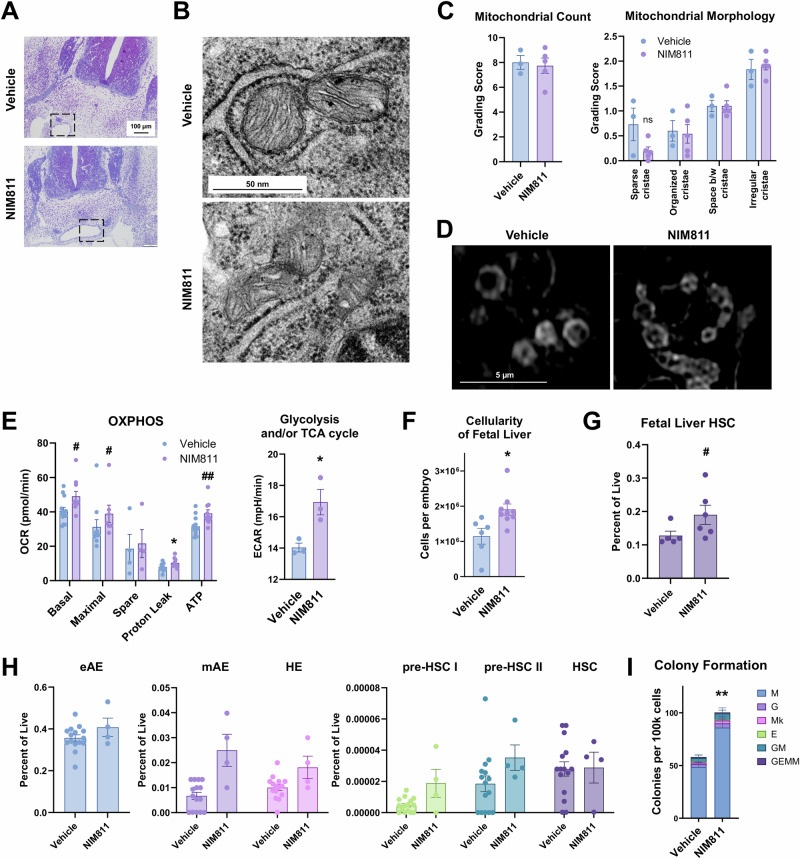
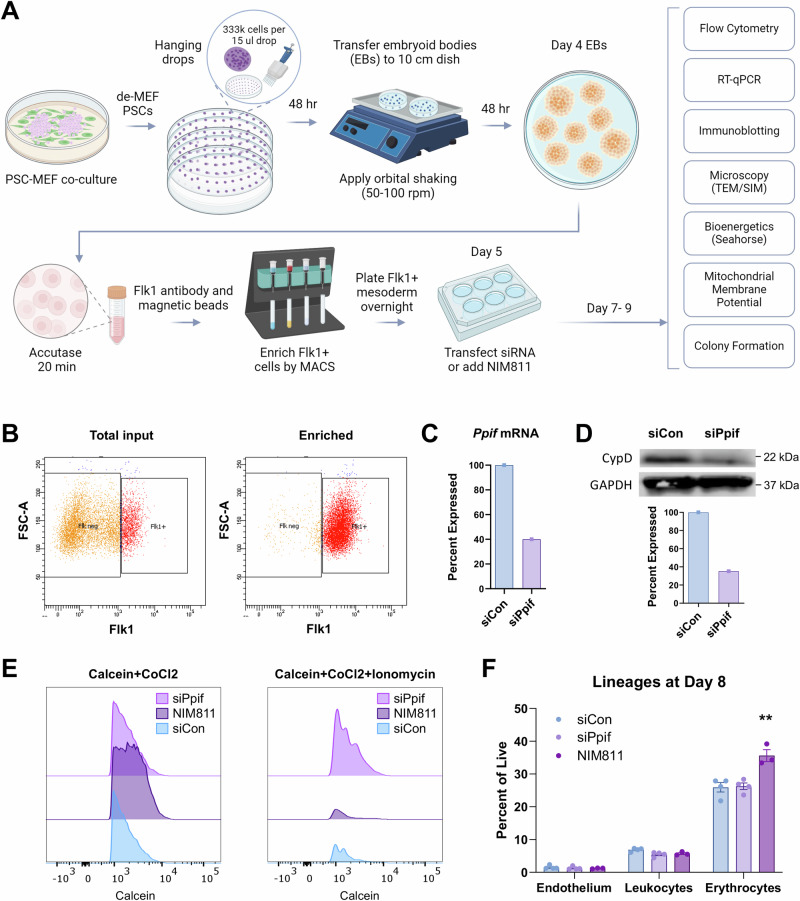
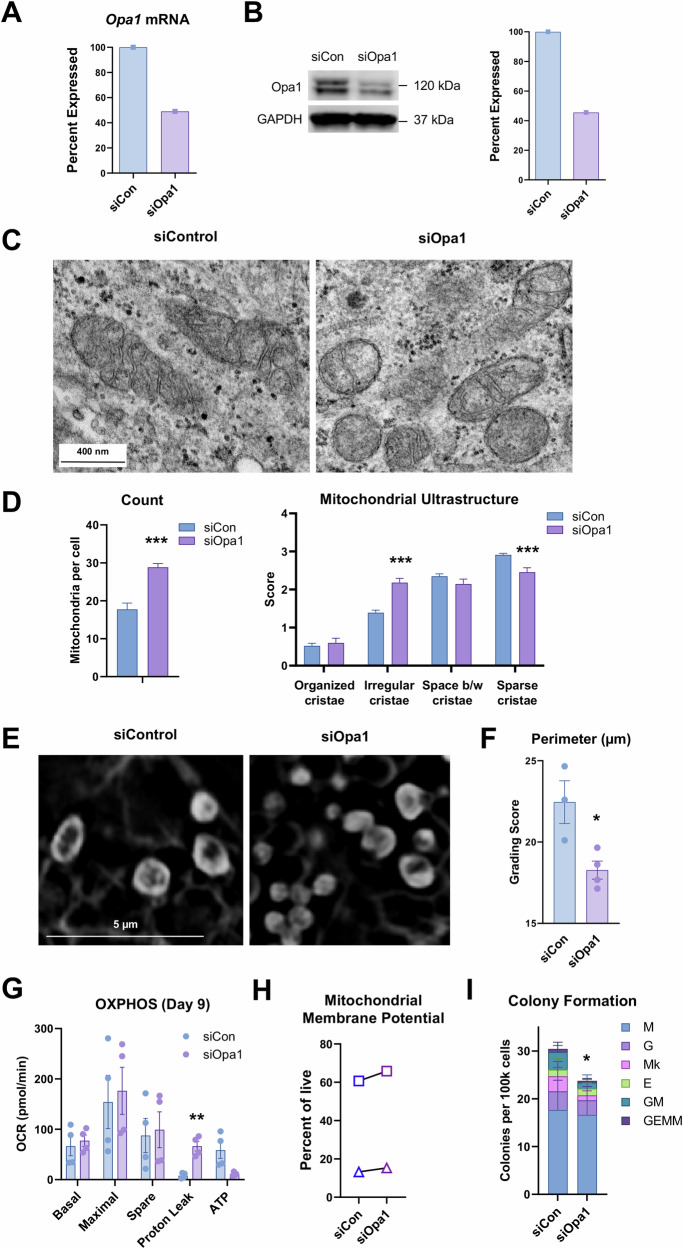
The mitochondrial permeability transition pore (mPTP) is a supramolecular channel that regulates exchange of solutes across cristae membranes, with executive roles in mitochondrial function and cell death. The contribution of the mPTP to normal physiology remains debated, although evidence implicates the mPTP in mitochondrial inner membrane remodeling in differentiating progenitor cells. Here, we demonstrate that strict control over mPTP conductance shapes metabolic machinery as cells transit toward hematopoietic identity. Cells undergoing the endothelial-to-hematopoietic transition (EHT) tightly control chief regulatory elements of the mPTP. During EHT, maturing arterial endothelium restricts mPTP activity just prior to hematopoietic commitment. After transition in cellular identity, mPTP conductance is restored. In utero treatment with NIM811, a molecule that blocks sensitization of the mPTP to opening by Cyclophilin D (CypD), amplifies oxidative phosphorylation (OXPHOS) in hematopoietic precursors and increases hematopoiesis in the embryo. Additionally, differentiating pluripotent stem cells (PSCs) acquire greater organization of mitochondrial cristae and hematopoietic activity following knockdown of the CypD gene, Ppif. Conversely, knockdown of Opa1, a GTPase critical for proper cristae architecture, induces cristae irregularity and impairs hematopoiesis. These data elucidate a mechanism that regulates mitochondrial maturation in hematopoietic precursors and underscore a role for the mPTP in the acquisition of hematopoietic fate.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
- K01DK092365/U.S. Department of Health & Human Services | NIH | National Institute of Diabetes and Digestive and Kidney Diseases (National Institute of Diabetes & Digestive & Kidney Diseases)
- R01DK111599/U.S. Department of Health & Human Services | NIH | National Institute of Diabetes and Digestive and Kidney Diseases (National Institute of Diabetes & Digestive & Kidney Diseases)
- K01 HL143111/HL/NHLBI NIH HHS/United States
- Scholar Award/American Society of Hematology (ASH)
- RP110776/Cancer Prevention and Research Institute of Texas (Cancer Prevention Research Institute of Texas)
LinkOut - more resources
Full Text Sources
Medical
