

Single-nucleus chromatin accessibility and transcriptomic map of breast tissues of women of diverse genetic ancestry
- PMID: 39122969
- PMCID: PMC11976273
- DOI: 10.1038/s41591-024-03011-9
Single-nucleus chromatin accessibility and transcriptomic map of breast tissues of women of diverse genetic ancestry
Erratum in
-
Author Correction: Single-nucleus chromatin accessibility and transcriptomic map of breast tissues of women of diverse genetic ancestry.Nat Med. 2025 May;31(5):1714. doi: 10.1038/s41591-025-03681-z. Nat Med. 2025. PMID: 40195453 Free PMC article. No abstract available.
Abstract
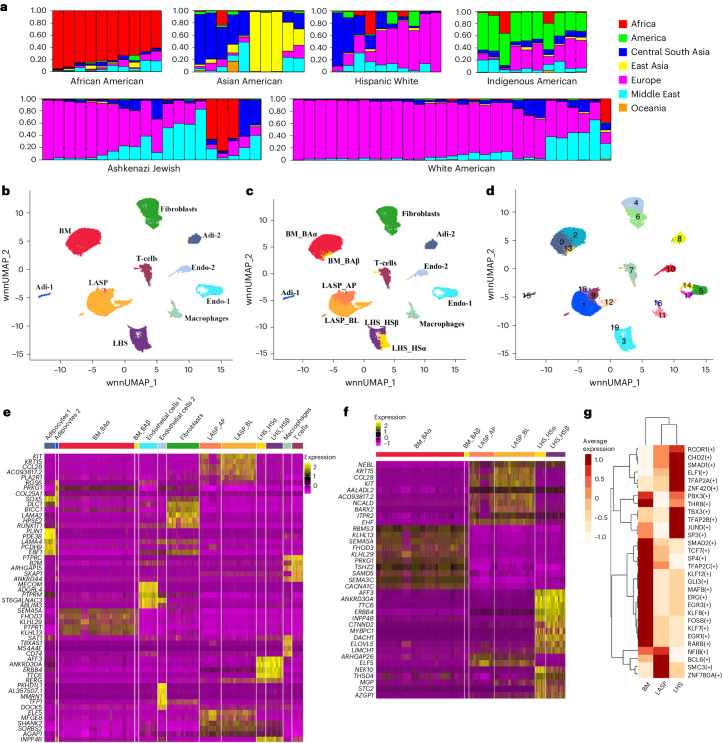
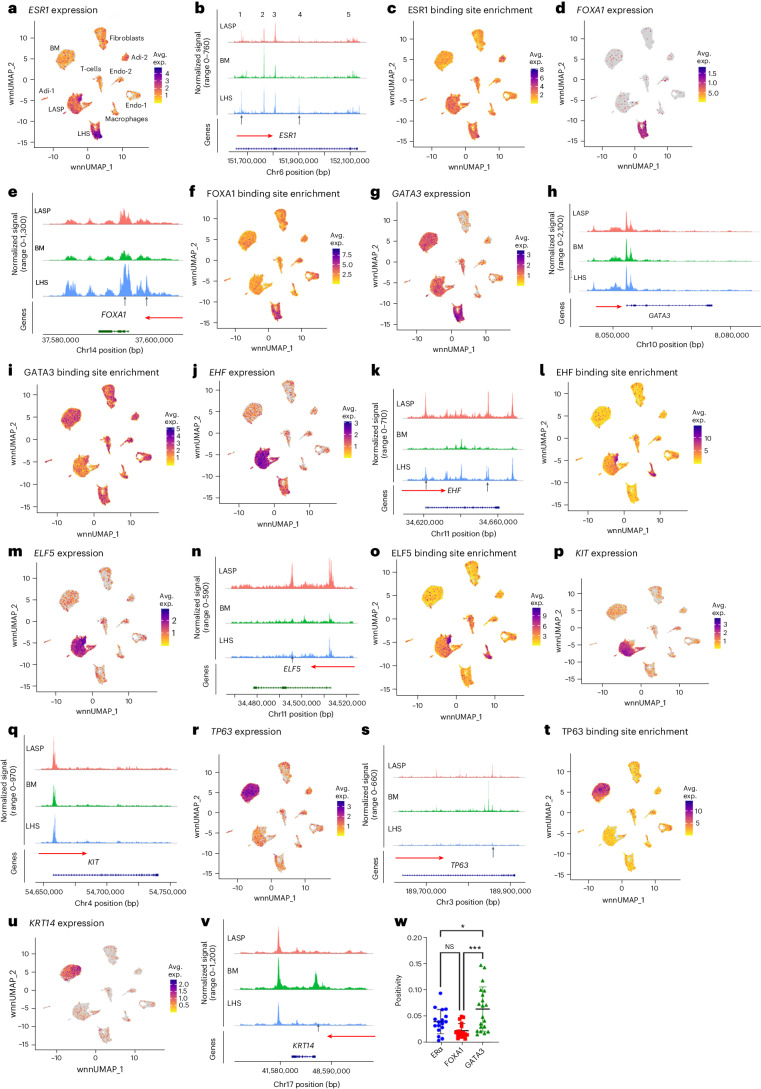
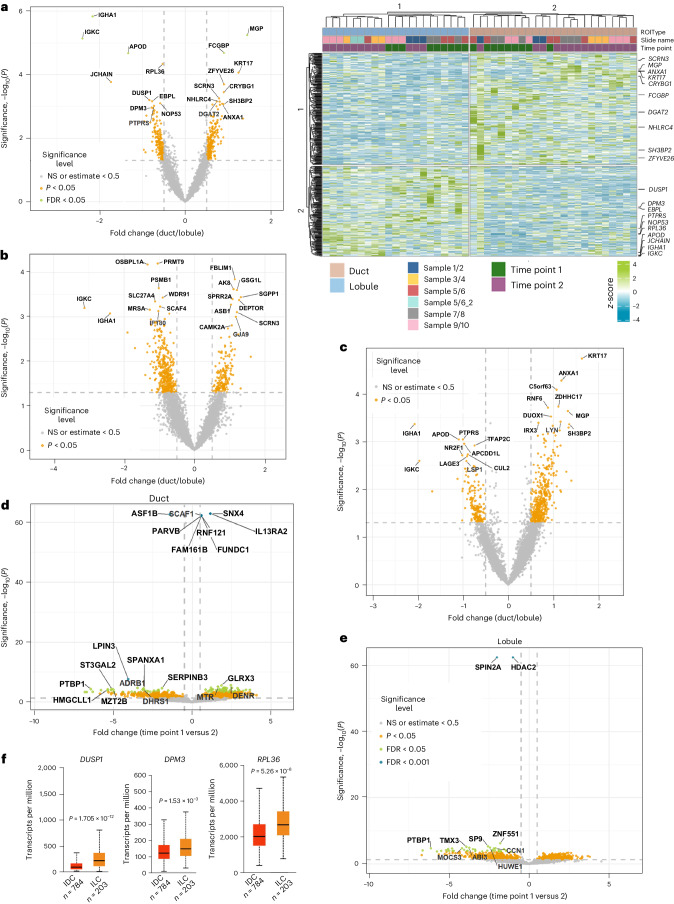
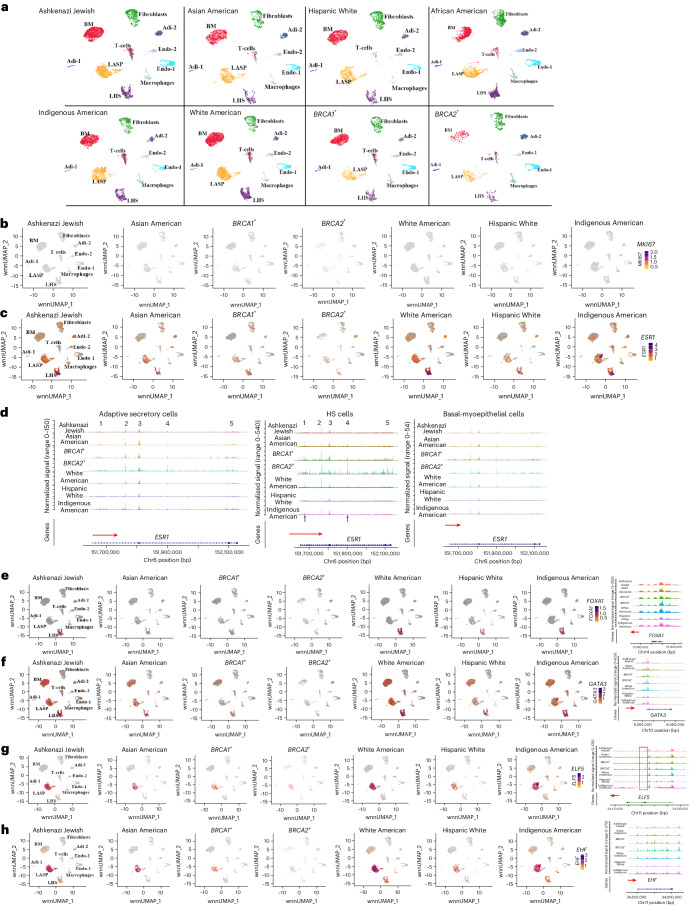
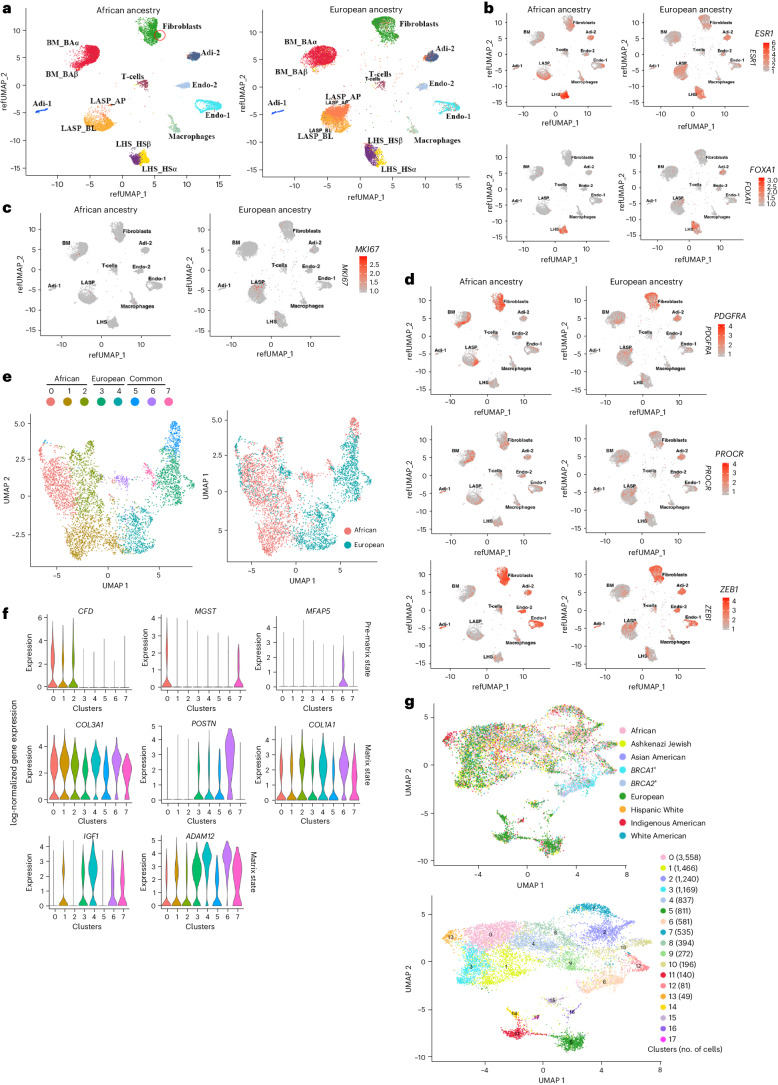
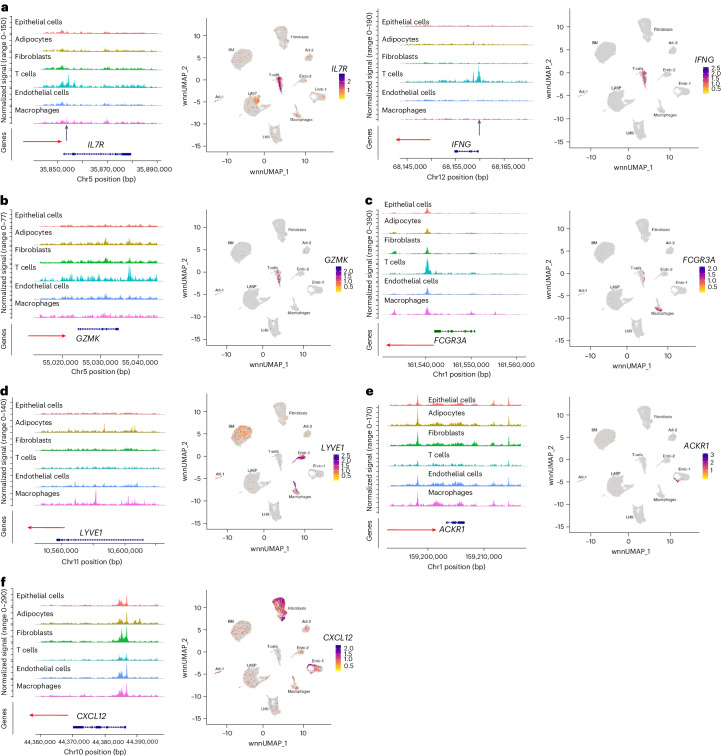
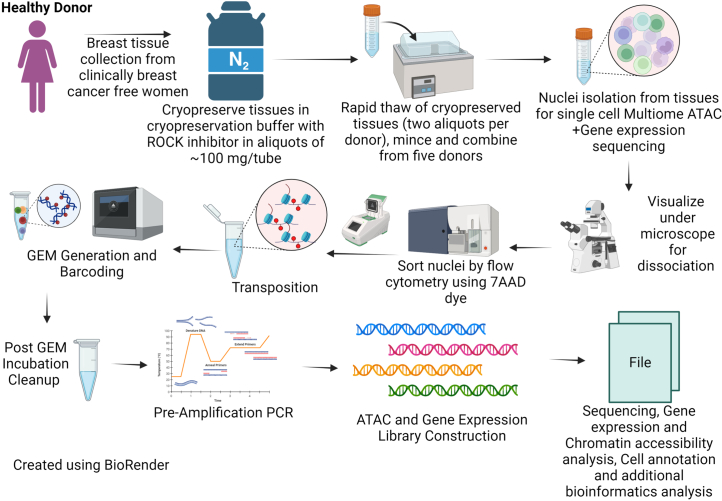
Single-nucleus analysis allows robust cell-type classification and helps to establish relationships between chromatin accessibility and cell-type-specific gene expression. Here, using samples from 92 women of several genetic ancestries, we developed a comprehensive chromatin accessibility and gene expression atlas of the breast tissue. Integrated analysis revealed ten distinct cell types, including three major epithelial subtypes (luminal hormone sensing, luminal adaptive secretory precursor (LASP) and basal-myoepithelial), two endothelial and adipocyte subtypes, fibroblasts, T cells, and macrophages. In addition to the known cell identity genes FOXA1 (luminal hormone sensing), EHF and ELF5 (LASP), TP63 and KRT14 (basal-myoepithelial), epithelial subtypes displayed several uncharacterized markers and inferred gene regulatory networks. By integrating breast epithelial cell gene expression signatures with spatial transcriptomics, we identified gene expression and signaling differences between lobular and ductal epithelial cells and age-associated changes in signaling networks. LASP cells and fibroblasts showed genetic ancestry-dependent variability. An estrogen receptor-positive subpopulation of LASP cells with alveolar progenitor cell state was enriched in women of Indigenous American ancestry. Fibroblasts from breast tissues of women of African and European ancestry clustered differently, with accompanying gene expression differences. Collectively, these data provide a vital resource for further exploring genetic ancestry-dependent variability in healthy breast biology.
© 2024. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Competing interests: The authors declare no competing interests.
Figures








References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
