

This is a preprint.
Synergy and antagonism in the integration of BCR and CD40 signals that control B-cell proliferation
- PMID: 39131345
- PMCID: PMC11312454
- DOI: 10.1101/2024.07.28.605521
Synergy and antagonism in the integration of BCR and CD40 signals that control B-cell proliferation
Update in
-
Synergy and antagonism in the integration of BCR and CD40 signals that control B-cell population expansion.Mol Syst Biol. 2025 Aug;21(8):1119-1146. doi: 10.1038/s44320-025-00124-2. Epub 2025 Jun 5. Mol Syst Biol. 2025. PMID: 40473841 Free PMC article.
Abstract
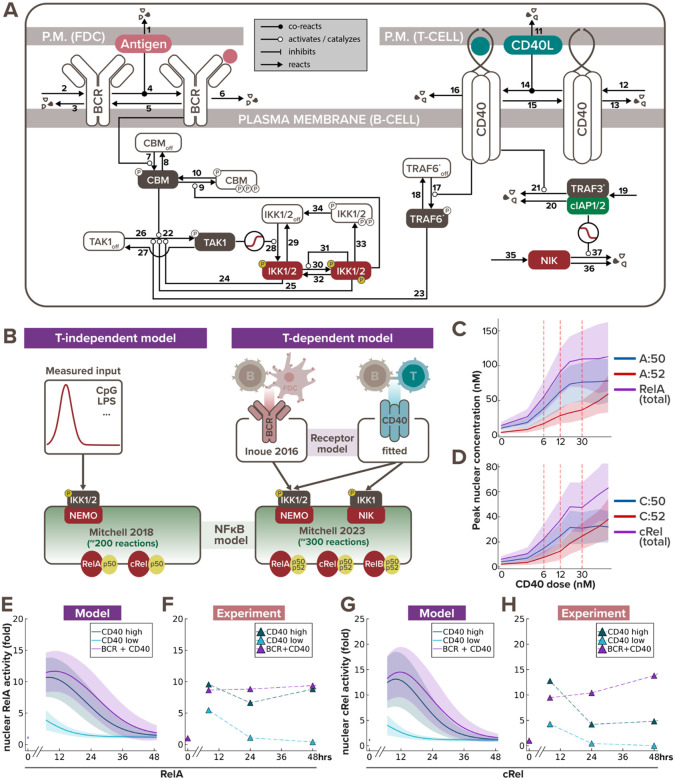
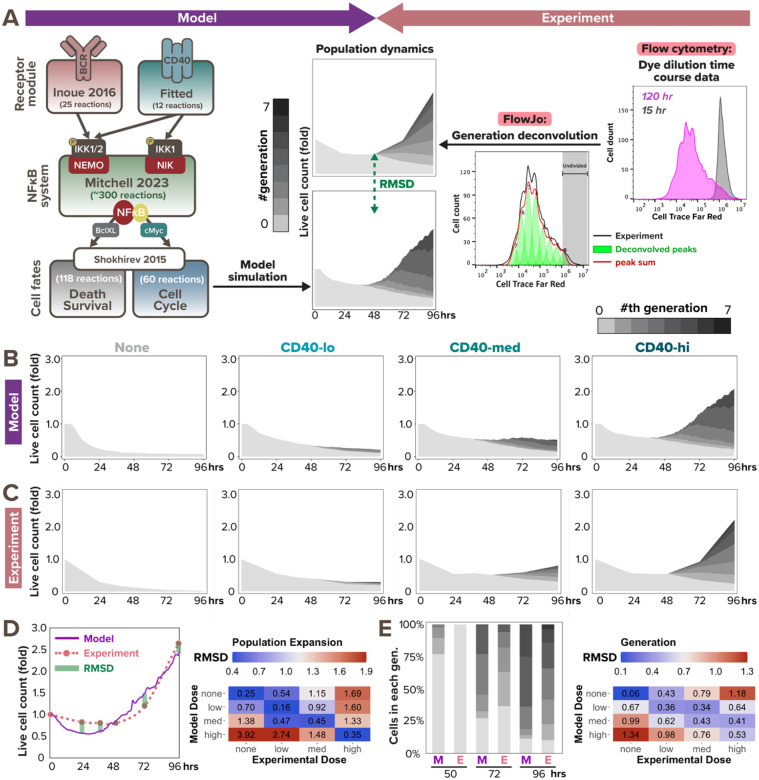
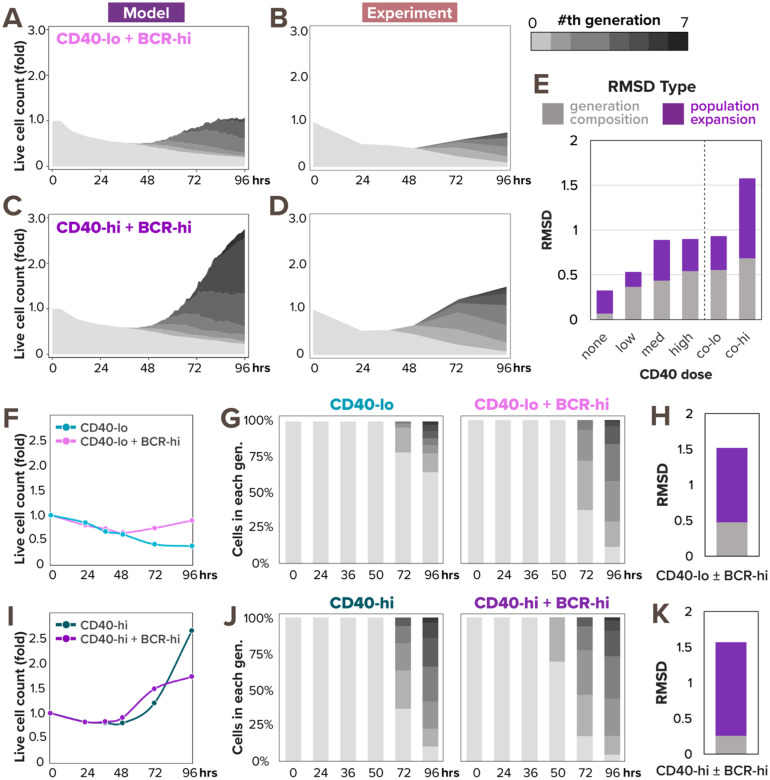
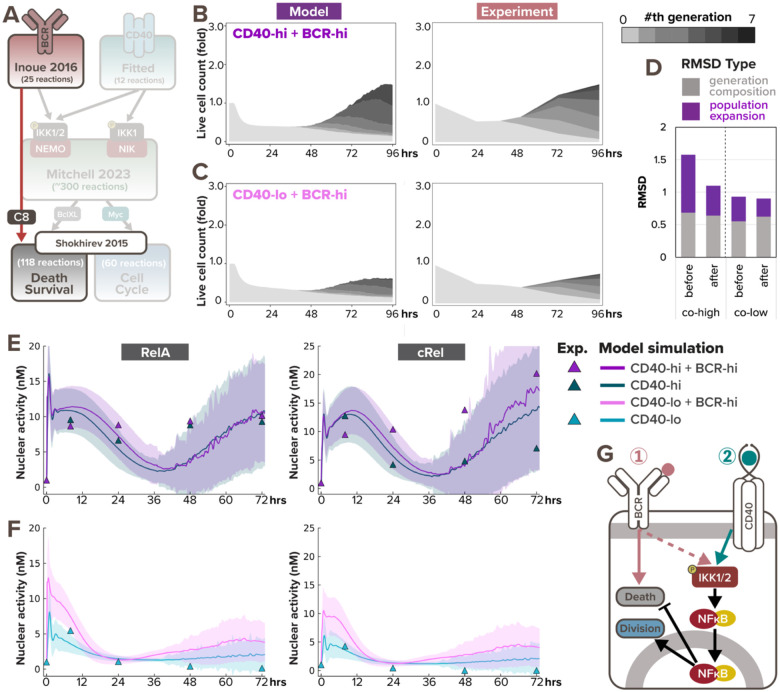
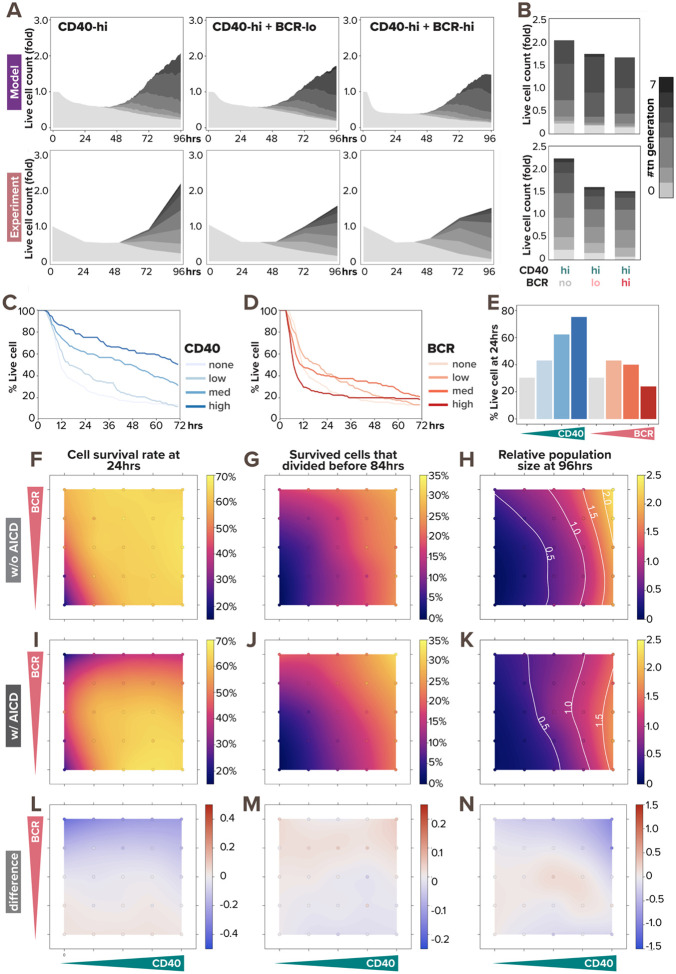
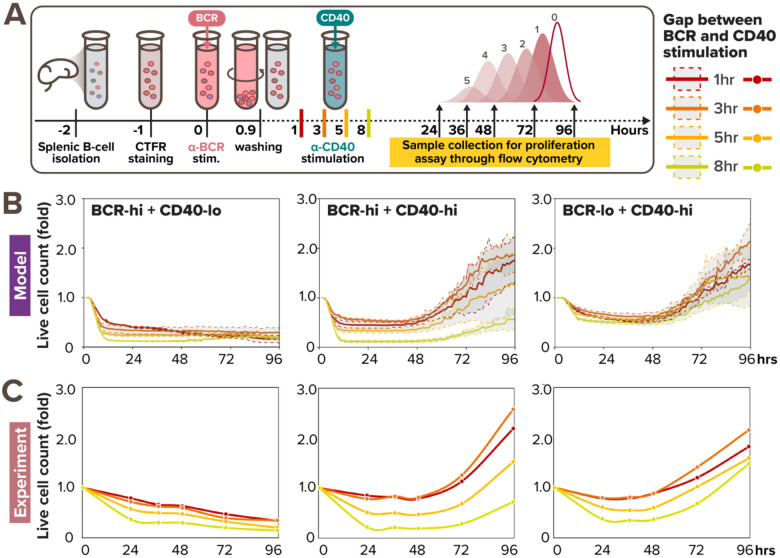
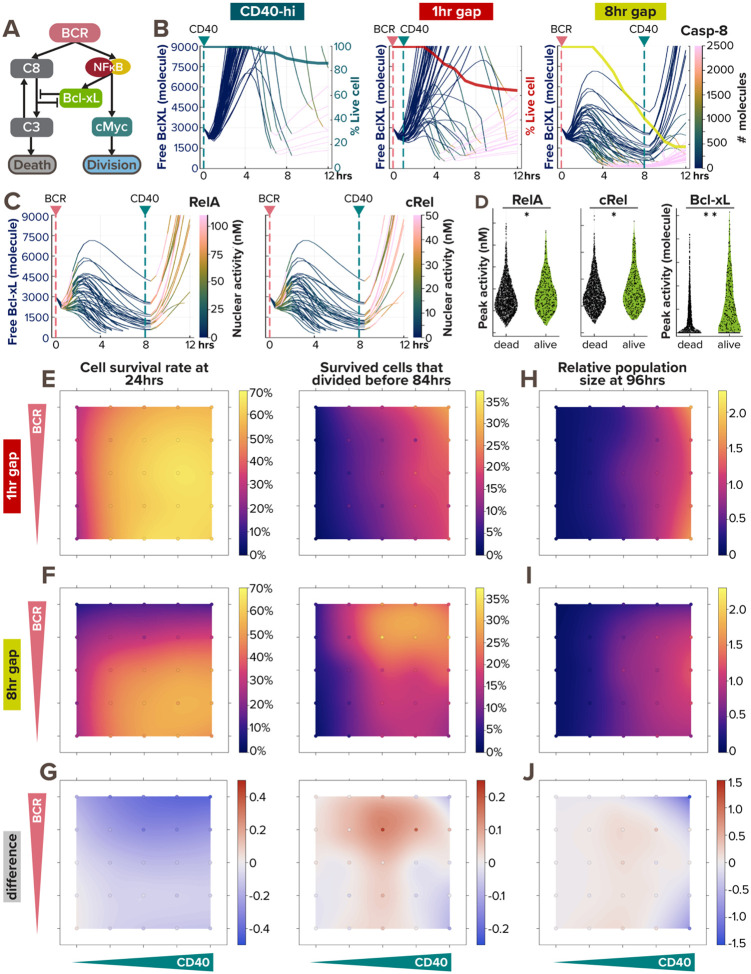
In response to infection or vaccination, a successful antibody response must enrich high-affinity antigen-reactive B-cells through positive selection, but eliminate auto-reactive B-cells through negative selection. B-cells receive signals from the B-cell receptor (BCR) which binds the antigen, and the CD40 receptor which is stimulated by neighboring T-cells that also recognize the antigen. How BCR and CD40 signaling are integrated quantitatively to jointly determine B-cell fate decision and proliferation remains unclear. To investigate this, we developed a differential-equations-based model of the BCR and CD40 signaling networks activating NFκB. Our model accurately recapitulates the NFκB dynamics of B-cells stimulated through their BCR and CD40 receptors, correctly predicting that costimulation induces more NFκB activity. However, when linking it to established cell fate decision models of cell survival and cell cycle control, it predicted potentiated population expansion that was not observed experimentally. We found that this discrepancy was due to a time-dependent functional antagonism exacerbated by BCR-induced caspase activity that can trigger apoptosis in founder cells, unless NFκB-induced survival gene expression protects B-cells in time. Guided by model predictions, sequential co-stimulation experiments revealed how the temporal dynamics of BCR and CD40 signaling control the fate decision between negative and positive selection of B-cell clonal expansion. Our quantitative findings highlight a complex non-monotonic integration of BCR and CD40 signals that is controlled by a balance between NFκB and cell-death pathways, and suggest a mechanism for regulating the stringency of B-cell selection during an antibody response.
Keywords: B-cell selection; BCR signaling; CD40 signaling; Germinal center reaction; NFκB; activation-induced cell death; mathematical modeling; systems immunology.
Conflict of interest statement
Conflict of Interest Statement The authors declare no conflict of interest.
Figures
References
-
- Bretscher P. and Cohn M. (1970) ‘A Theory of Self-Nonself Discrimination’, Science, 169(3950), pp. 1042–1049. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials