

Porphyromonas gingivalis fuels colorectal cancer through CHI3L1-mediated iNKT cell-driven immune evasion
- PMID: 39132842
- PMCID: PMC11321422
- DOI: 10.1080/19490976.2024.2388801
Porphyromonas gingivalis fuels colorectal cancer through CHI3L1-mediated iNKT cell-driven immune evasion
Abstract
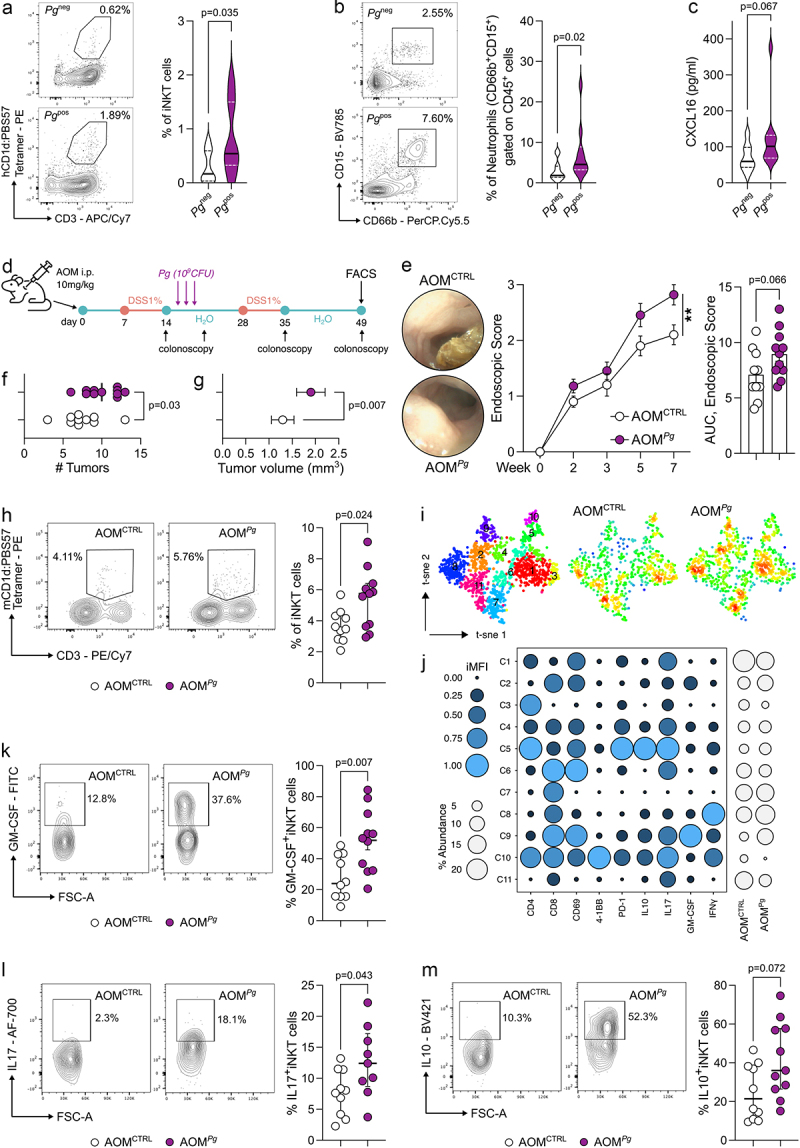
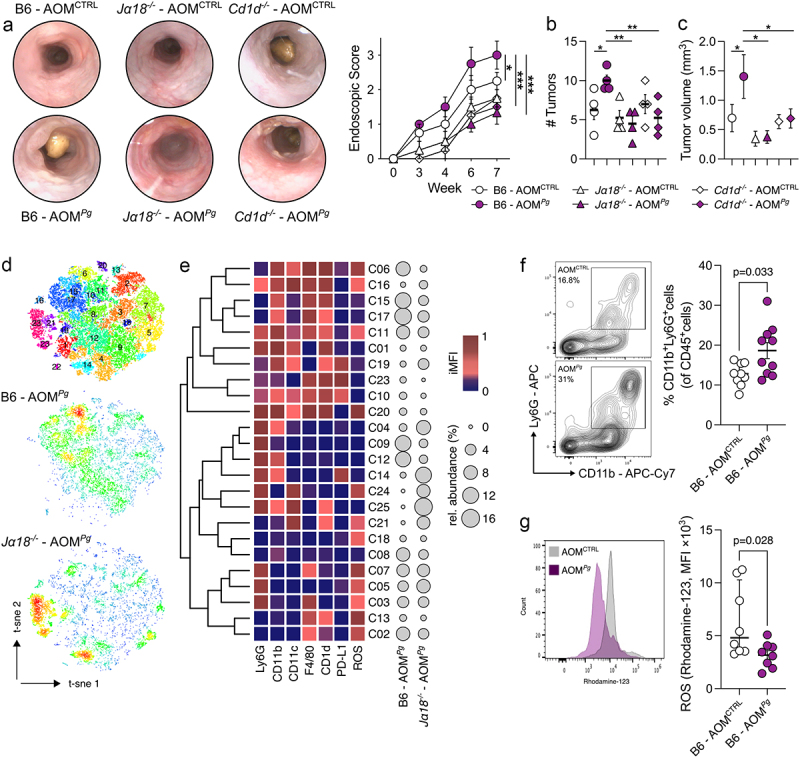
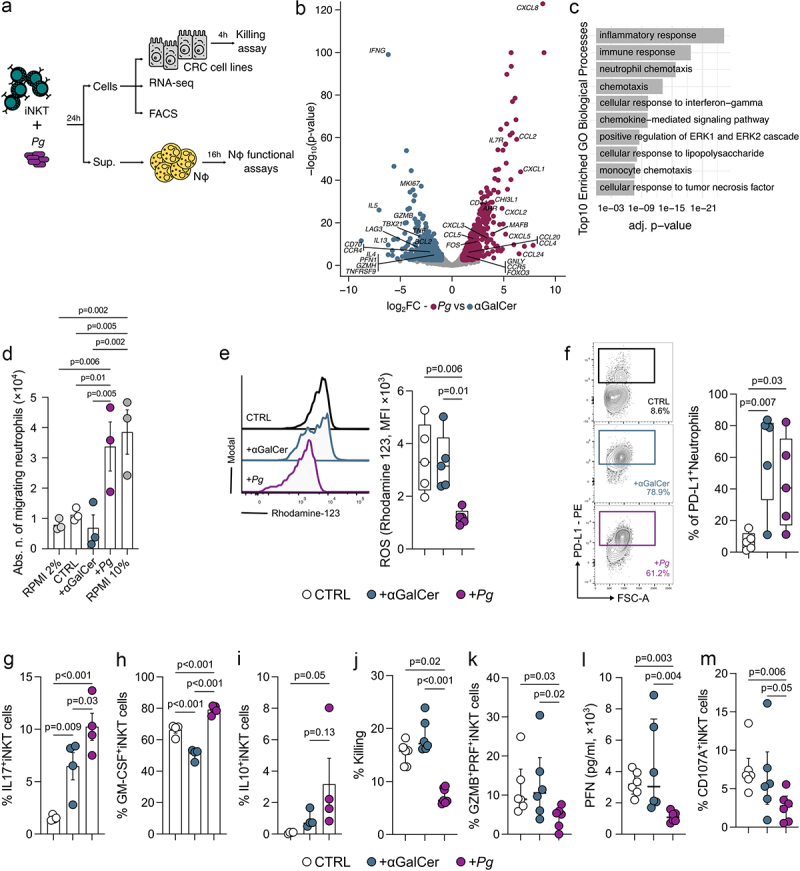
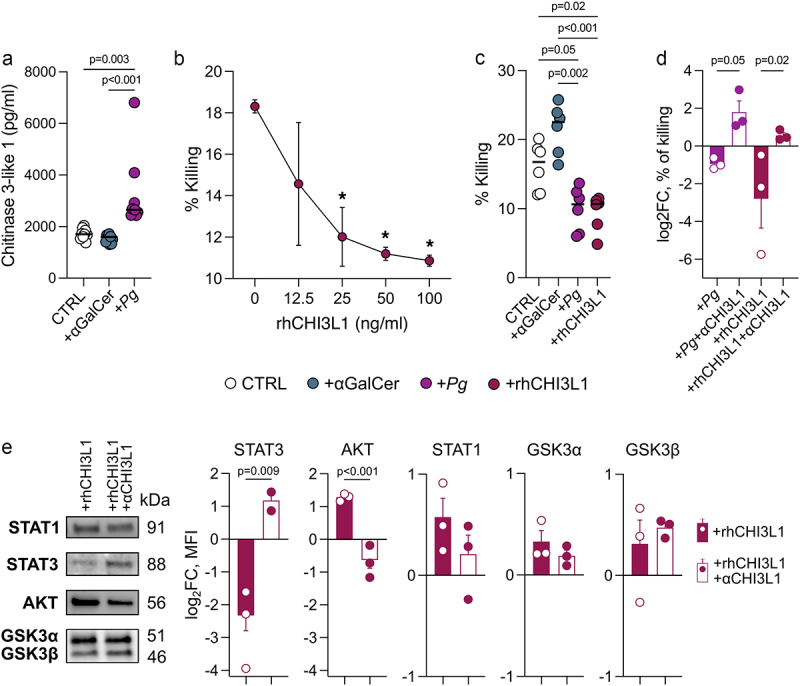
The interaction between the gut microbiota and invariant Natural Killer T (iNKT) cells plays a pivotal role in colorectal cancer (CRC). The pathobiont Fusobacterium nucleatum influences the anti-tumor functions of CRC-infiltrating iNKT cells. However, the impact of other bacteria associated with CRC, like Porphyromonas gingivalis, on their activation status remains unexplored. In this study, we demonstrate that mucosa-associated P. gingivalis induces a protumour phenotype in iNKT cells, subsequently influencing the composition of mononuclear-phagocyte cells within the tumor microenvironment. Mechanistically, in vivo and in vitro experiments showed that P. gingivalis reduces the cytotoxic functions of iNKT cells, hampering the iNKT cell lytic machinery through increased expression of chitinase 3-like-1 protein (CHI3L1). Neutralization of CHI3L1 effectively restores iNKT cell cytotoxic functions suggesting a therapeutic potential to reactivate iNKT cell-mediated antitumour immunity. In conclusion, our data demonstrate how P. gingivalis accelerates CRC progression by inducing the upregulation of CHI3L1 in iNKT cells, thus impairing their cytotoxic functions and promoting host tumor immune evasion.
Keywords: CHI3L1; CRC; Porphyromonas gingivalis; iNKT cells.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases