

Expansion of the HSV-2-specific T cell repertoire in skin after immunotherapeutic HSV-2 vaccine
- PMID: 39133650
- PMCID: PMC11383358
- DOI: 10.1172/jci.insight.179010
Expansion of the HSV-2-specific T cell repertoire in skin after immunotherapeutic HSV-2 vaccine
Abstract
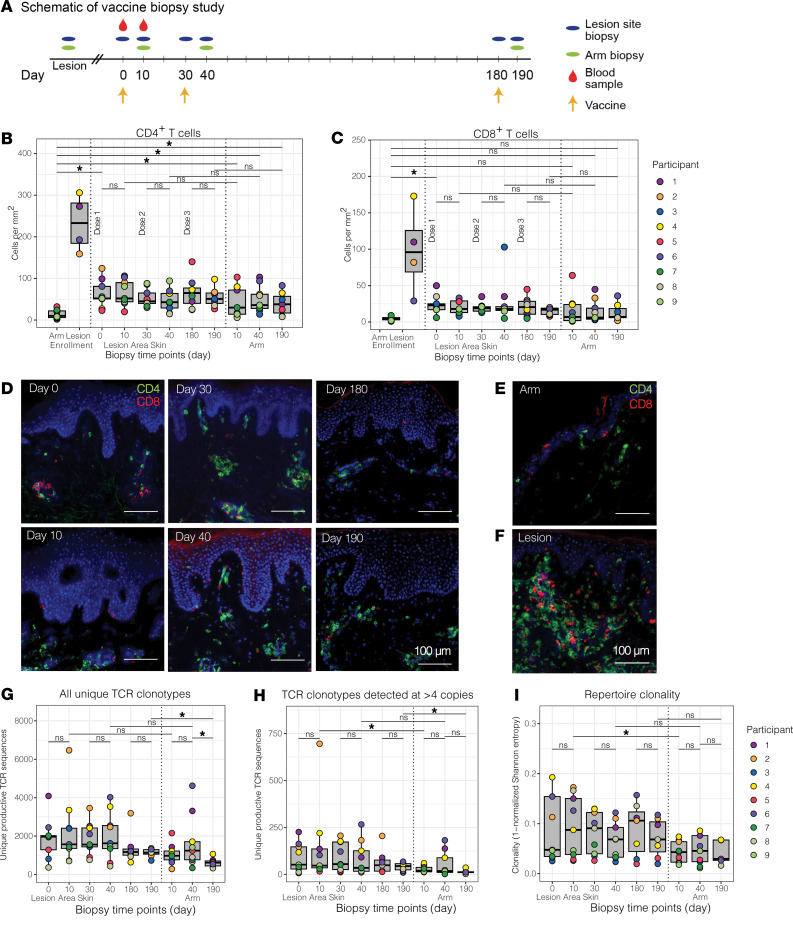
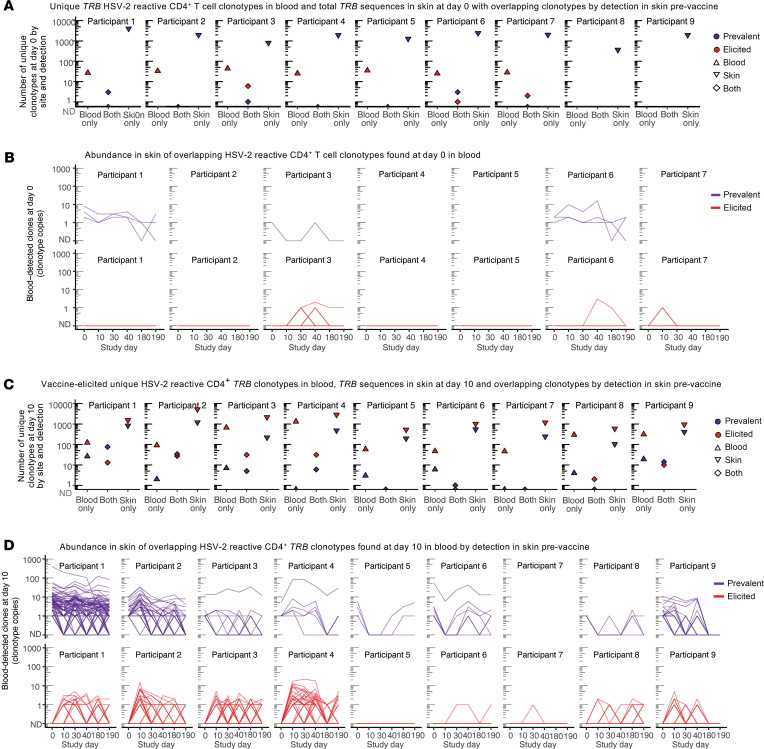
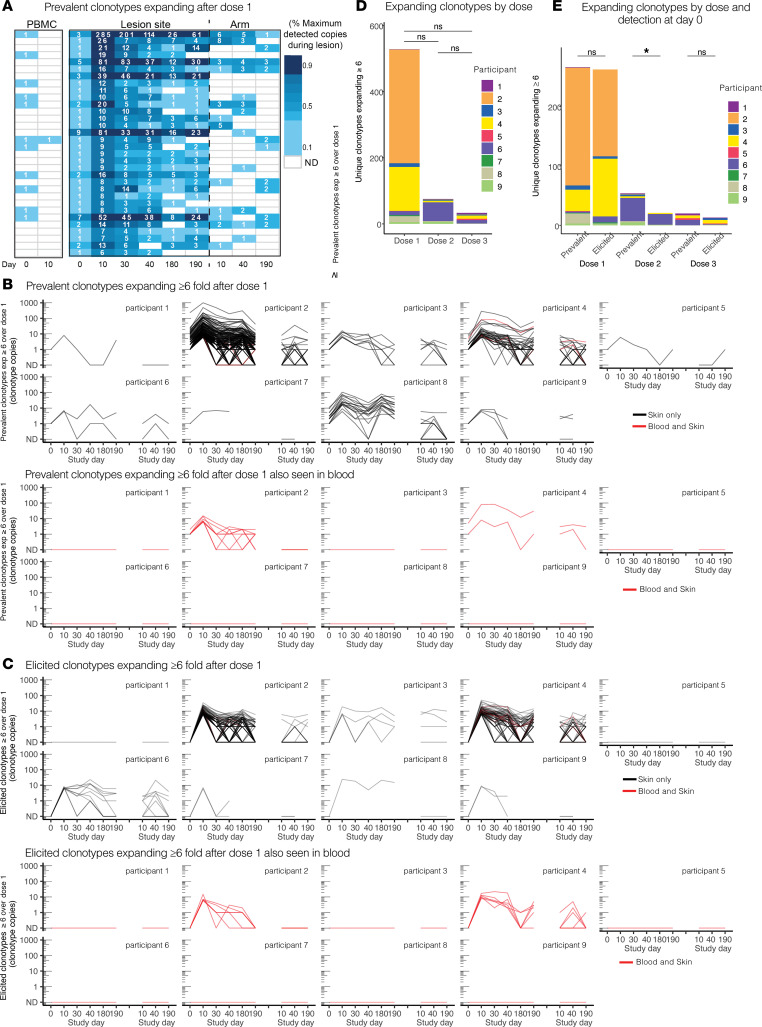
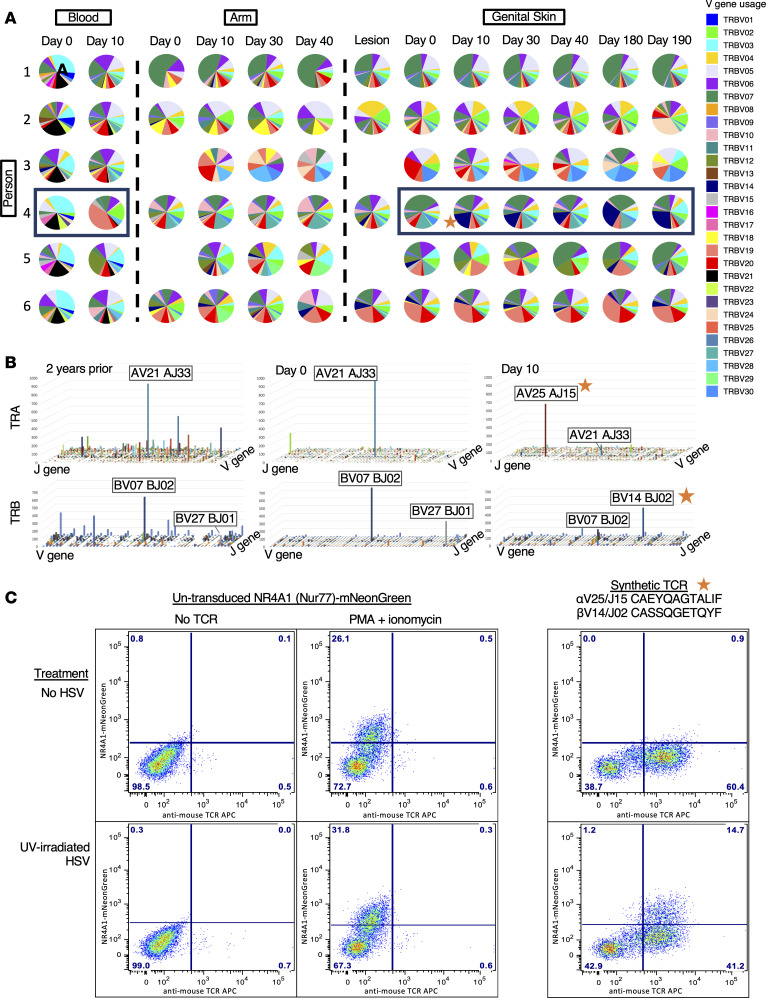
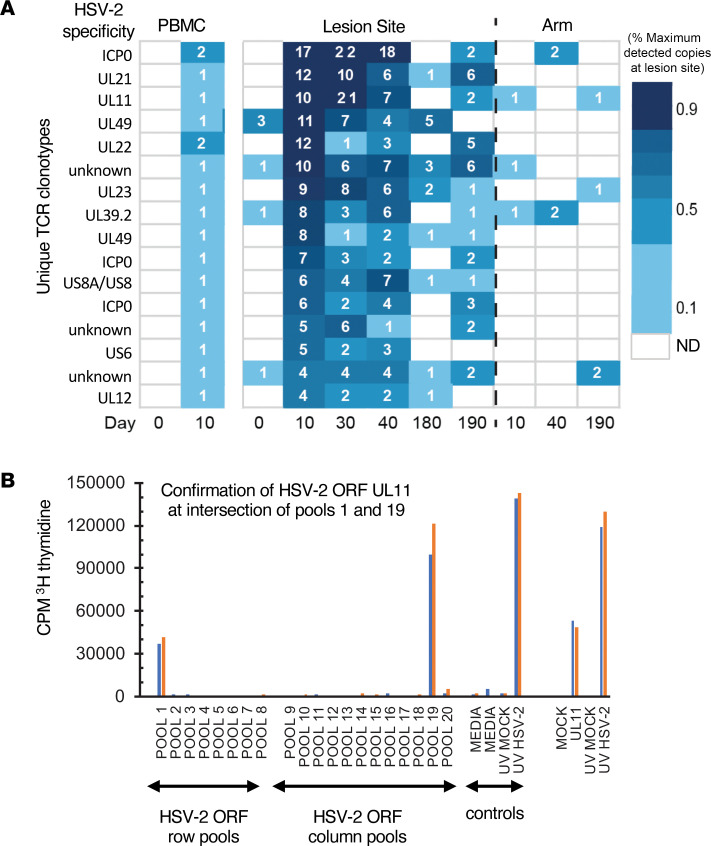
The skin at the site of HSV-2 reactivation is enriched for HSV-2-specific T cells. To evaluate whether an immunotherapeutic vaccine could elicit skin-based memory T cells, we studied skin biopsies and HSV-2-reactive CD4+ T cells from PBMCs by T cell receptor (TCR) β chain (TRB) sequencing before and after vaccination with a replication-incompetent whole-virus HSV-2 vaccine candidate (HSV529). The representation of HSV-2-reactive CD4+ TRB sequences from PBMCs in the skin TRB repertoire increased after the first vaccine dose. We found sustained expansion after vaccination of unique, skin-based T cell clonotypes that were not detected in HSV-2-reactive CD4+ T cells isolated from PBMCs. In one participant, a switch in immunodominance occurred with the emergence of a TCR αβ pair after vaccination that was not detected in blood. This TCRαβ was shown to be HSV-2 reactive by expression of a synthetic TCR in a Jurkat-based NR4A1 reporter system. The skin in areas of HSV-2 reactivation possessed an oligoclonal TRB repertoire that was distinct from the circulation. Defining the influence of therapeutic vaccination on the HSV-2-specific TRB repertoire requires tissue-based evaluation.
Keywords: Adaptive immunity; Cellular immune response; T cell receptor; Vaccines; Virology.
Conflict of interest statement
Figures
Update of
-
Expansion of the HSV-2-specific T cell repertoire in skin after immunotherapeutic HSV-2 vaccine.medRxiv [Preprint]. 2024 Feb 1:2022.02.04.22270210. doi: 10.1101/2022.02.04.22270210. medRxiv. 2024. Update in: JCI Insight. 2024 Jun 18;9(14):e179010. doi: 10.1172/jci.insight.179010. PMID: 38352384 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
