

MYB52 negatively regulates ADF9-meditated actin filament bundling in Arabidopsis pavement cell morphogenesis
- PMID: 39136601
- PMCID: PMC11583842
- DOI: 10.1111/jipb.13762
MYB52 negatively regulates ADF9-meditated actin filament bundling in Arabidopsis pavement cell morphogenesis
Abstract
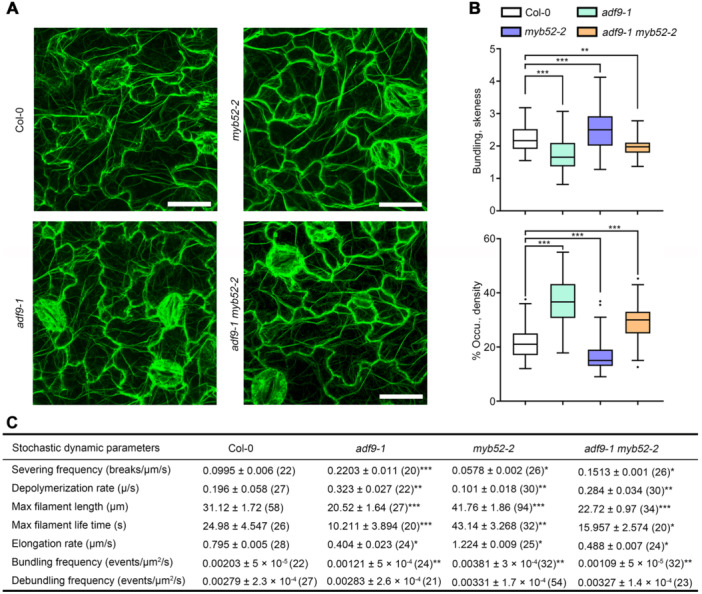
It has been proposed that cortical fine actin filaments are needed for the morphogenesis of pavement cells (PCs). However, the precise role and regulation mechanisms of actin filaments in PC morphogenesis are not well understood. Here, we found that Arabidopsis thaliana ACTIN DEPOLYMERIZING FACTOR9 (ADF9) is required for the morphogenesis of PC, which is negatively regulated by the R2R3 MYELOBLASTOSIS (MYB) transcription factor MYB52. In adf9 mutants, the lobe number of cotyledon PCs was significantly reduced, while the average lobe length did not differ significantly compared to that of wild type (Col-0), except for the variations in cell area and circularity, whereas the PC shapes in ADF9 overexpression seedlings showed different results. ADF9 decorated actin filaments, and colocalized with plasma membrane. The extent of filament bundling and actin filament bundling activity in adf9 mutant decreased. In addition, MYB52 directly targeted the promoter of ADF9 and negatively regulated its expression. The myb52-2 mutant showed increased lobe number and cell area, reduced cell circularity of PCs, and the PC phenotypes were suppressed when ADF9 was knocked out. Taken together, our data demonstrate that actin filaments play an important role in the morphogenesis of PC and reveal a transcriptional mechanism underlying MYB52 regulation of ADF9-mediated actin filament bundling in PC morphogenesis.
Keywords: ADF9; MYB52; actin filament bundling; pavement cell.
© 2024 The Author(s). Journal of Integrative Plant Biology published by John Wiley & Sons Australia, Ltd on behalf of Institute of Botany, Chinese Academy of Sciences.
Conflict of interest statement
The authors declare no conflict of interest.
Figures








References
-
- Altartouri, B. , Bidhendi, A.J. , Tani, T. , Suzuki, J. , Conrad, C. , Chebli, Y. , Liu, N. , Karunakaran, C. , Scarcelli, G. , and Geitmann, A. (2019). Pectin chemistry and cellulose crystallinity govern pavement cell morphogenesis in a multi‐step mechanism. Plant Physiol. 181: 127–141. - PMC - PubMed
-
- Andrianantoandro, E. , and Pollard, T.D. (2006). Mechanism of actin filament turnover by severing and nucleation at different concentrations of ADF/cofilin. Mol. Cell 24: 13–23. - PubMed
-
- Belteton, S.A. , Li, W. , Yanagisawa, M. , Hatam, F.A. , Quinn, M.I. , Szymanski, M.K. , Marley, M.W. , Turner, J.A. , and Szymanski, D.B. (2021). Real‐time conversion of tissue‐scale mechanical forces into an interdigitated growth pattern. Nat Plants 7: 826–841. - PubMed
-
- Bi, S. , Li, M. , Liu, C. , Liu, X. , Cheng, J. , Wang, L. , Wang, J. , Lv, Y. , He, M. , Cheng, X. , et al. (2022). Actin depolymerizing factor ADF7 inhibits actin bundling protein VILLIN1 to regulate root hair formation in response to osmotic stress in Arabidopsis. PLoS Genet. 18: e1010338. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
