

Multi-tiered chemical proteomic maps of tryptoline acrylamide-protein interactions in cancer cells
- PMID: 39138346
- PMCID: PMC11684312
- DOI: 10.1038/s41557-024-01601-1
Multi-tiered chemical proteomic maps of tryptoline acrylamide-protein interactions in cancer cells
Abstract
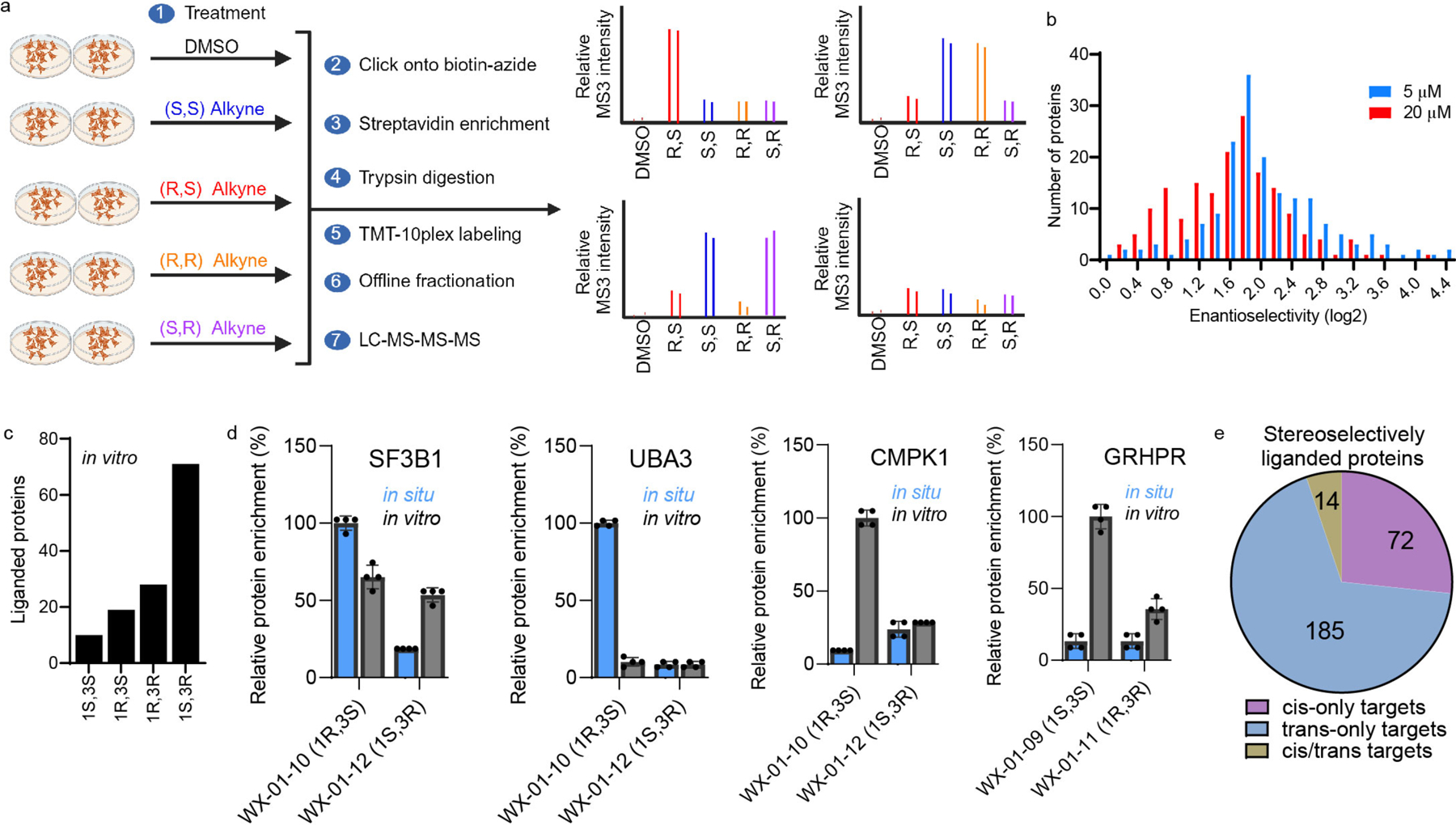
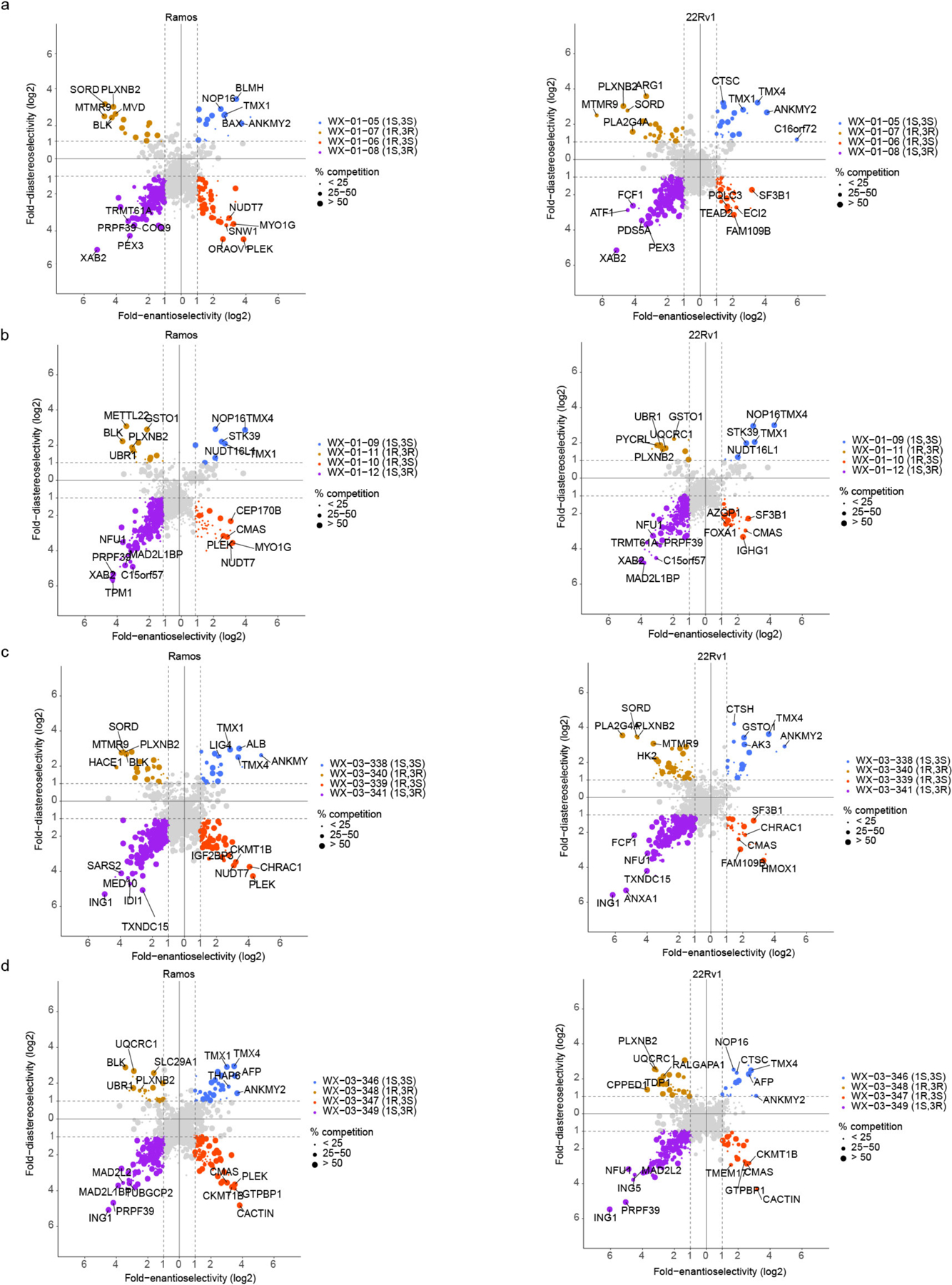
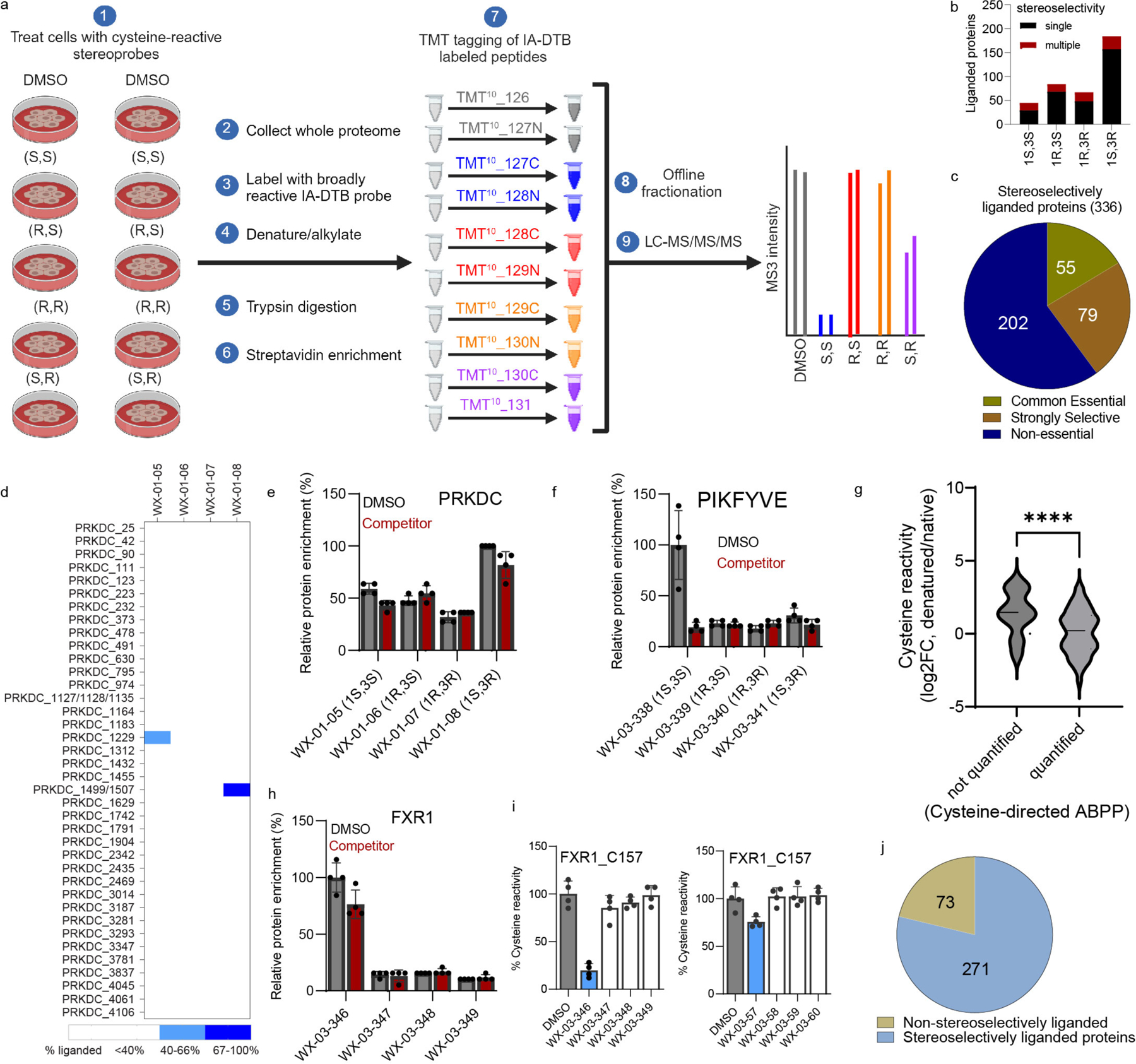
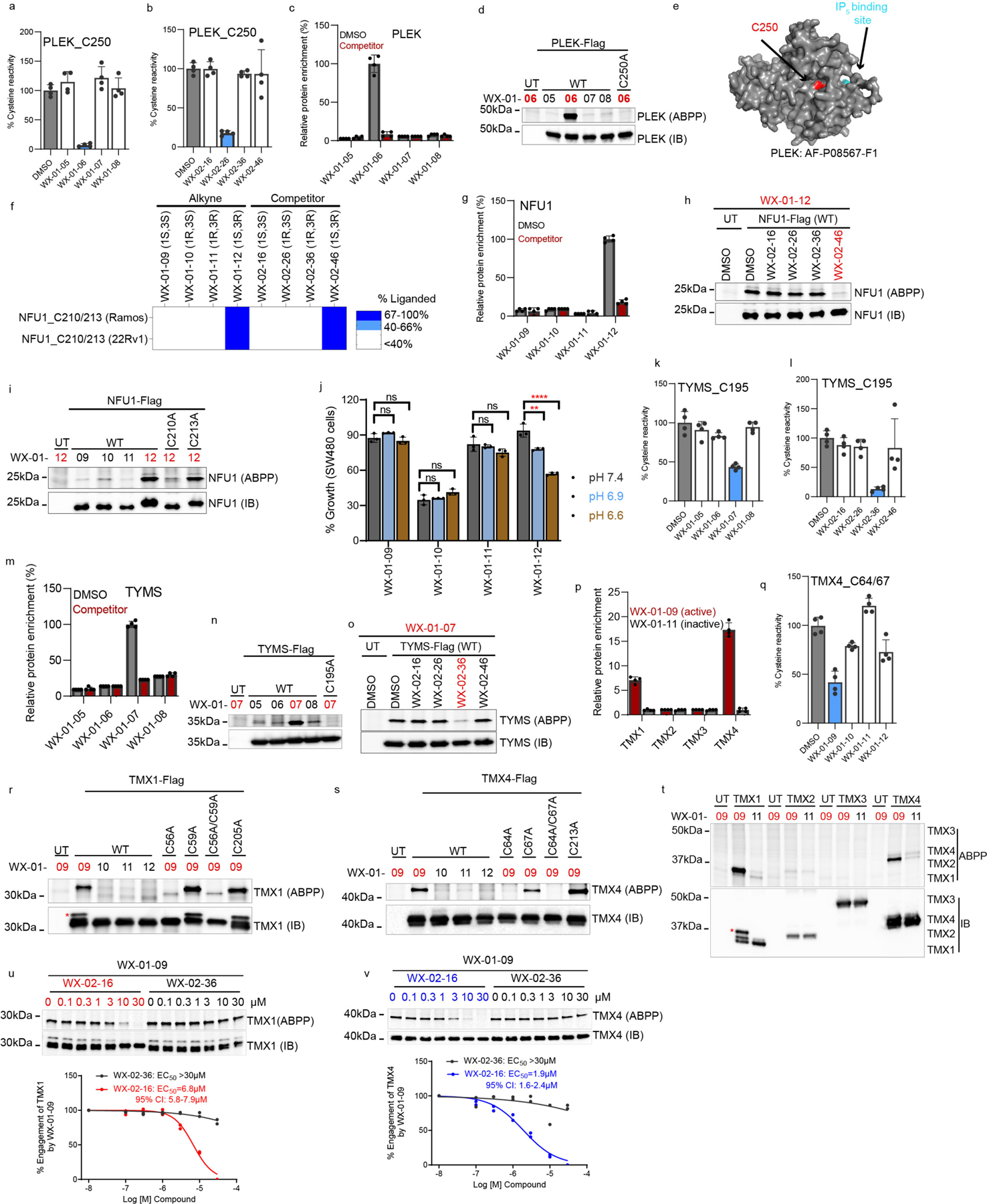
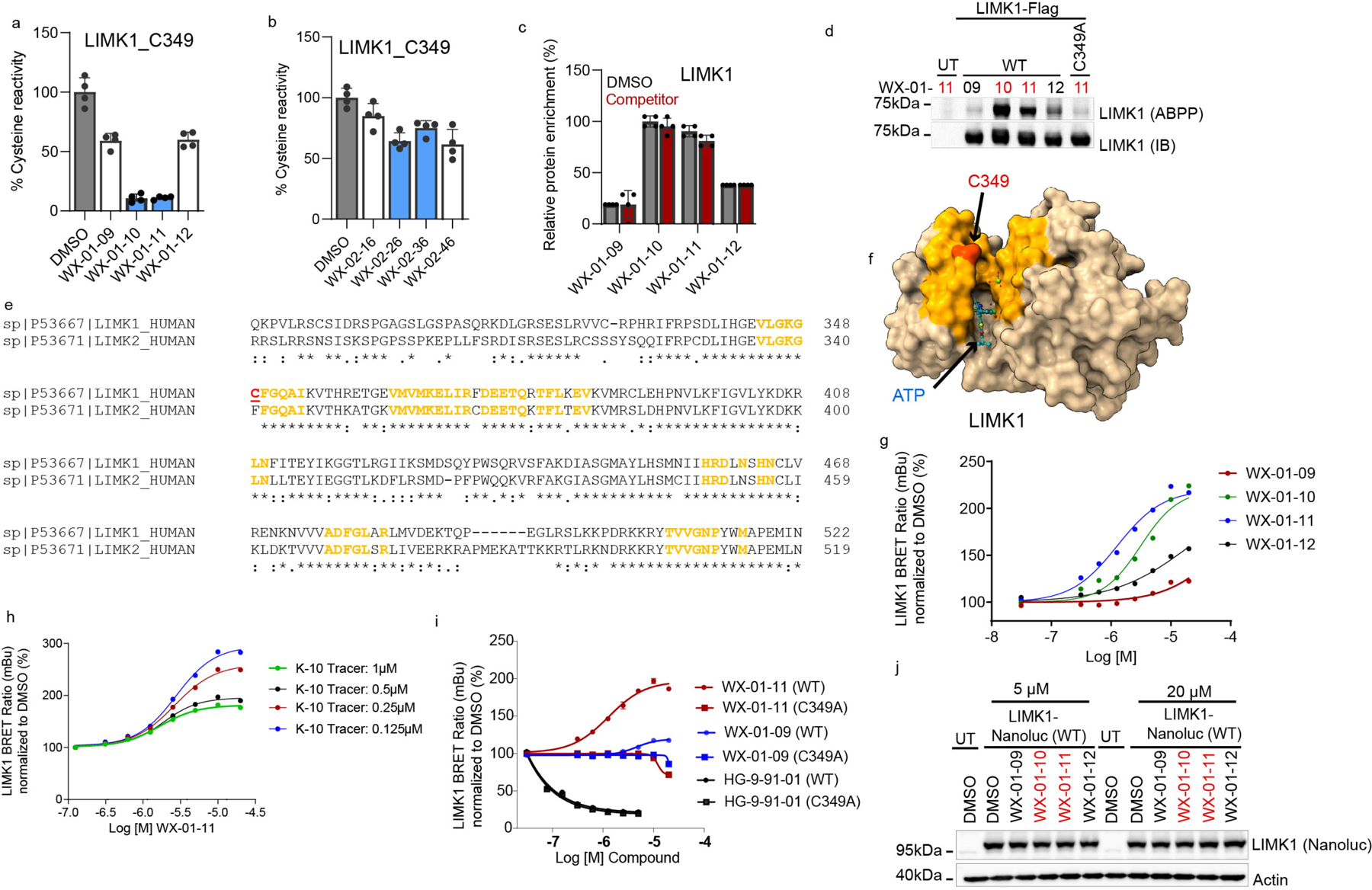
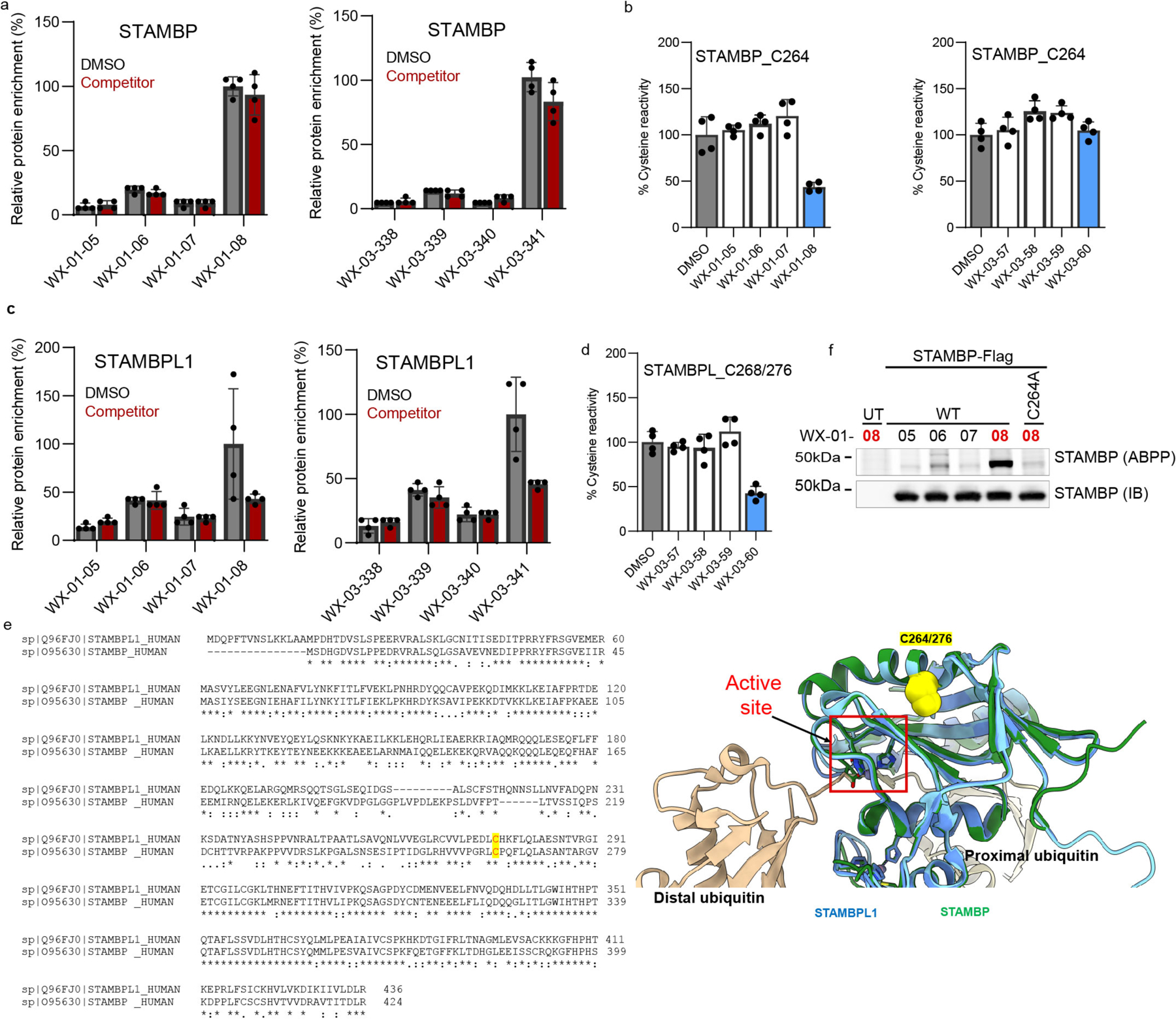
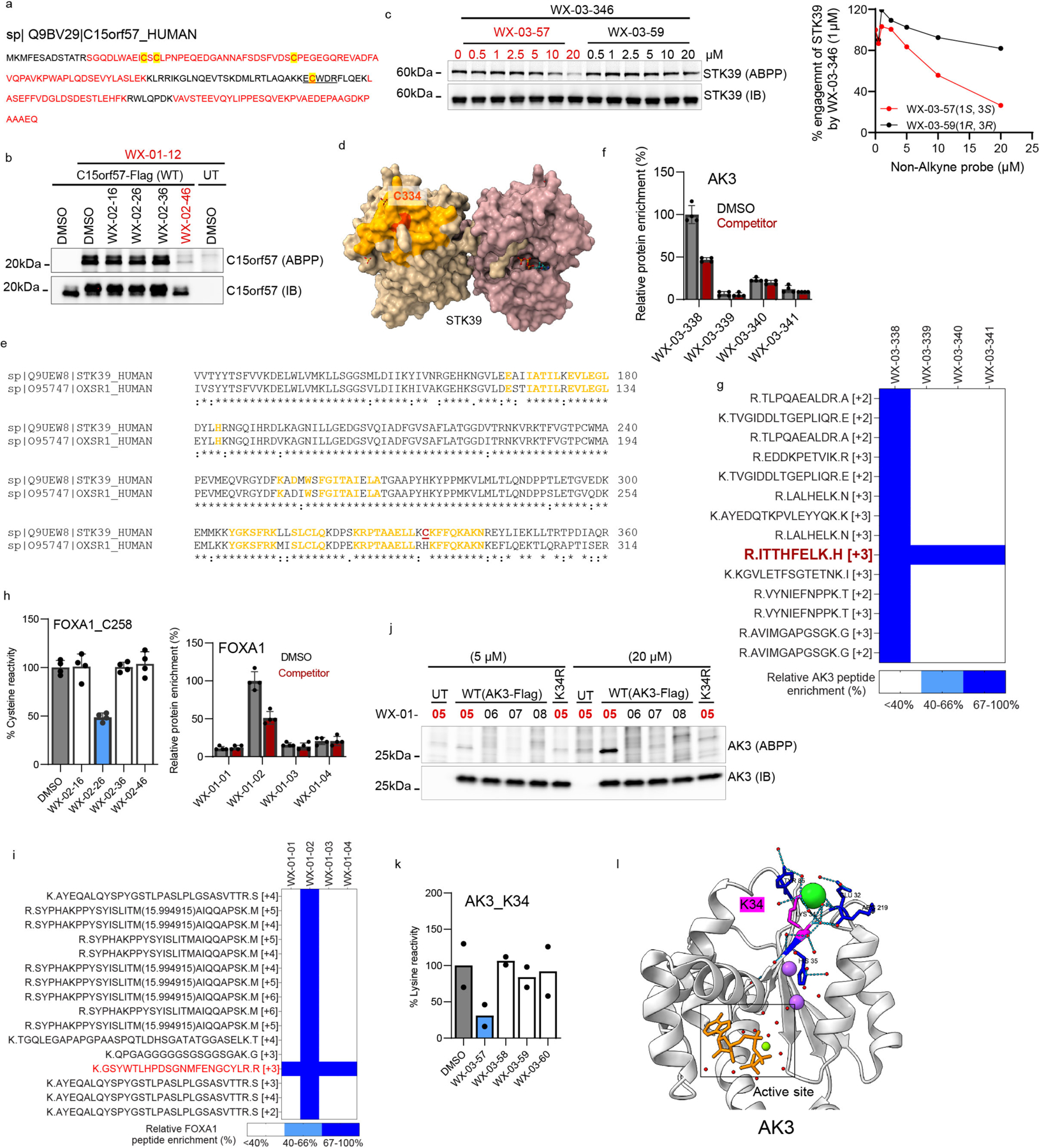
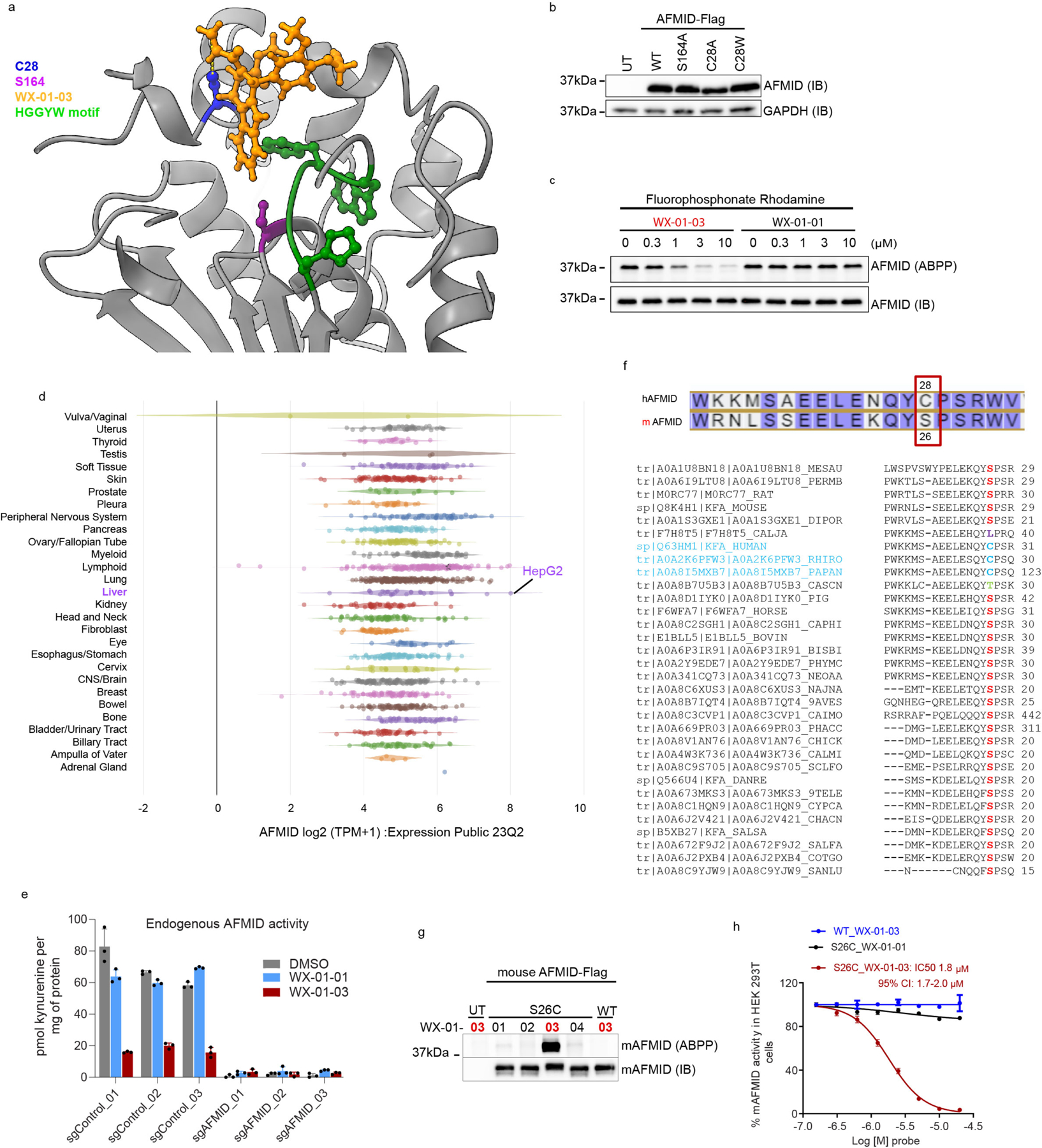
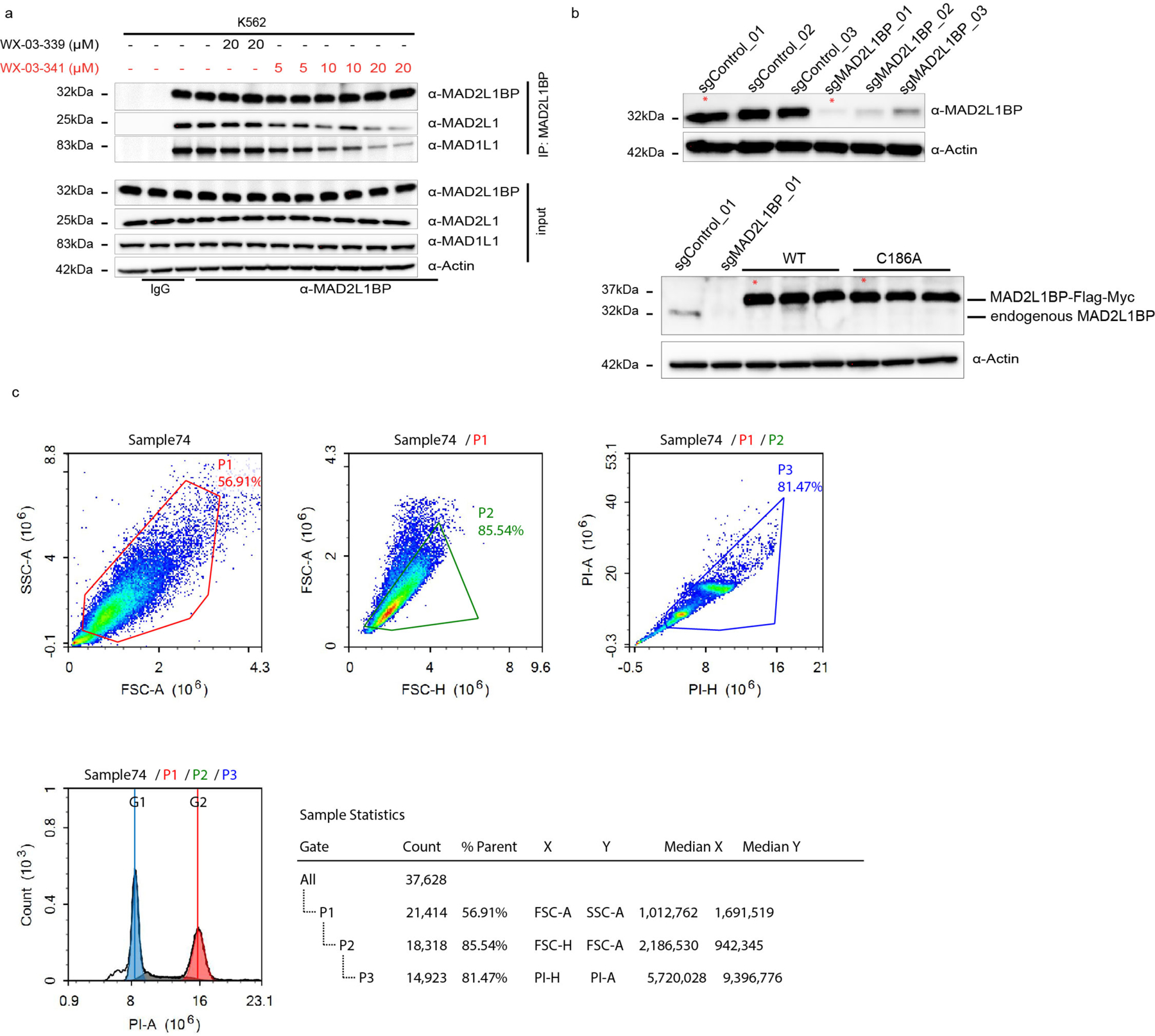
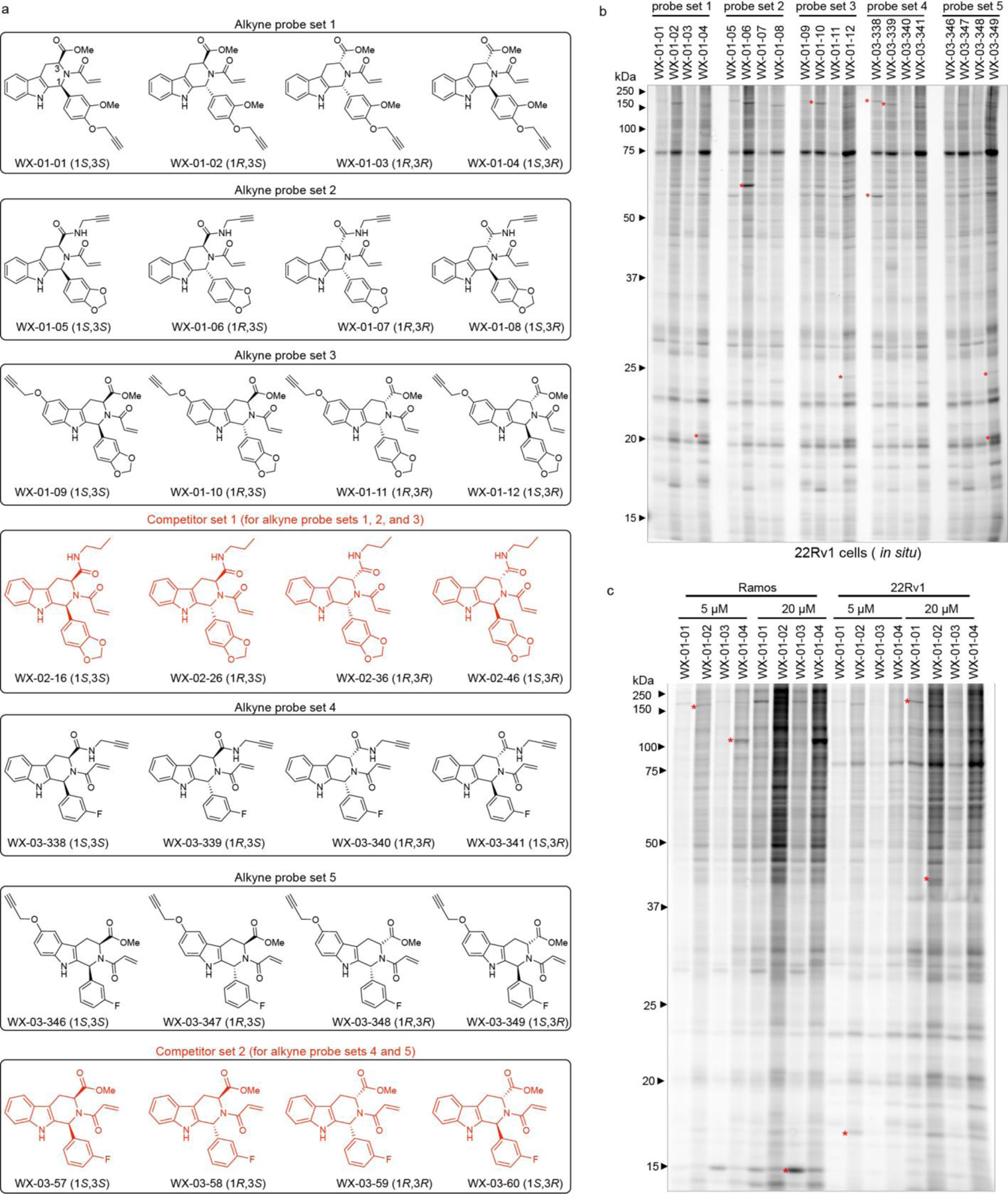
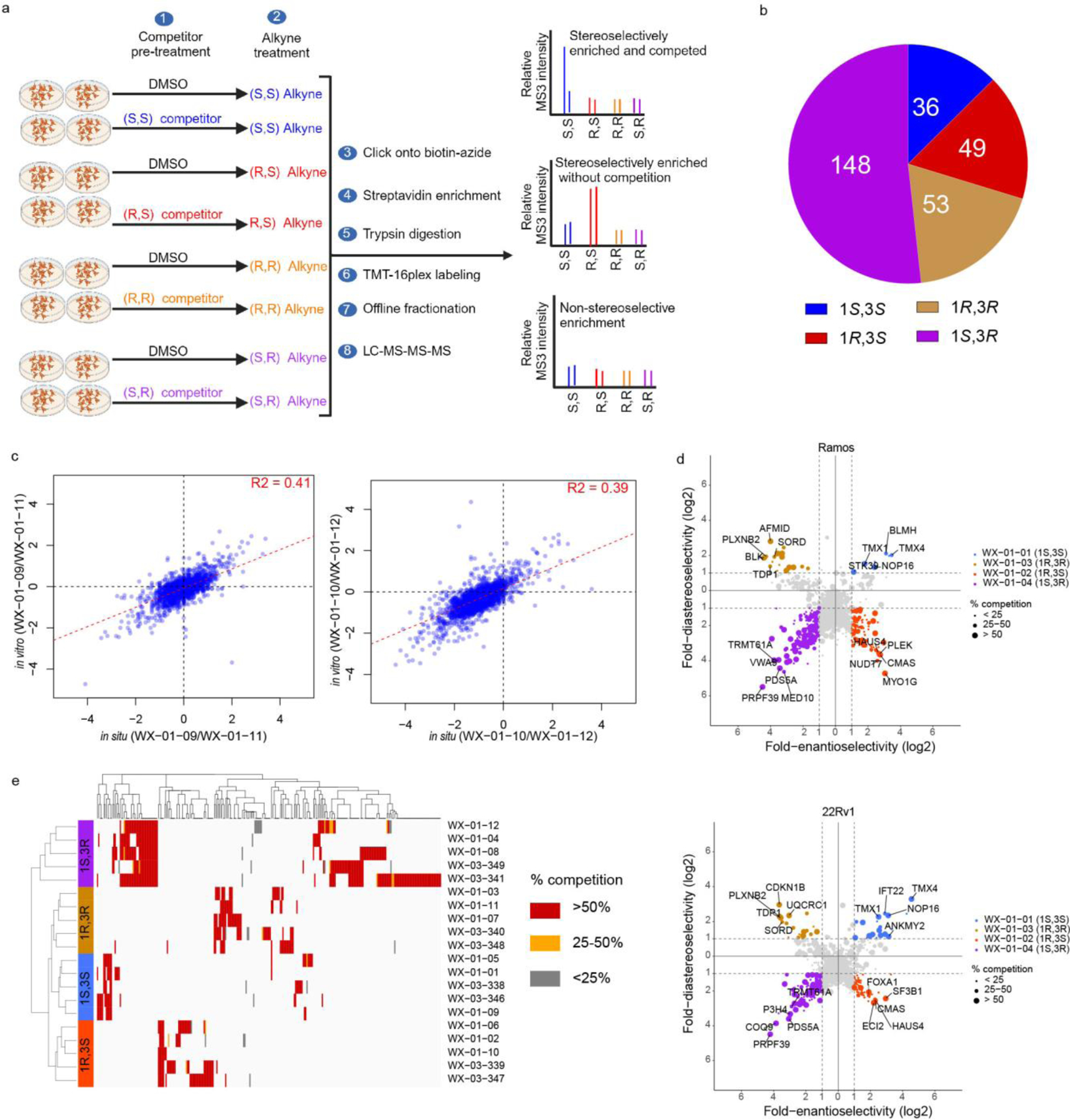
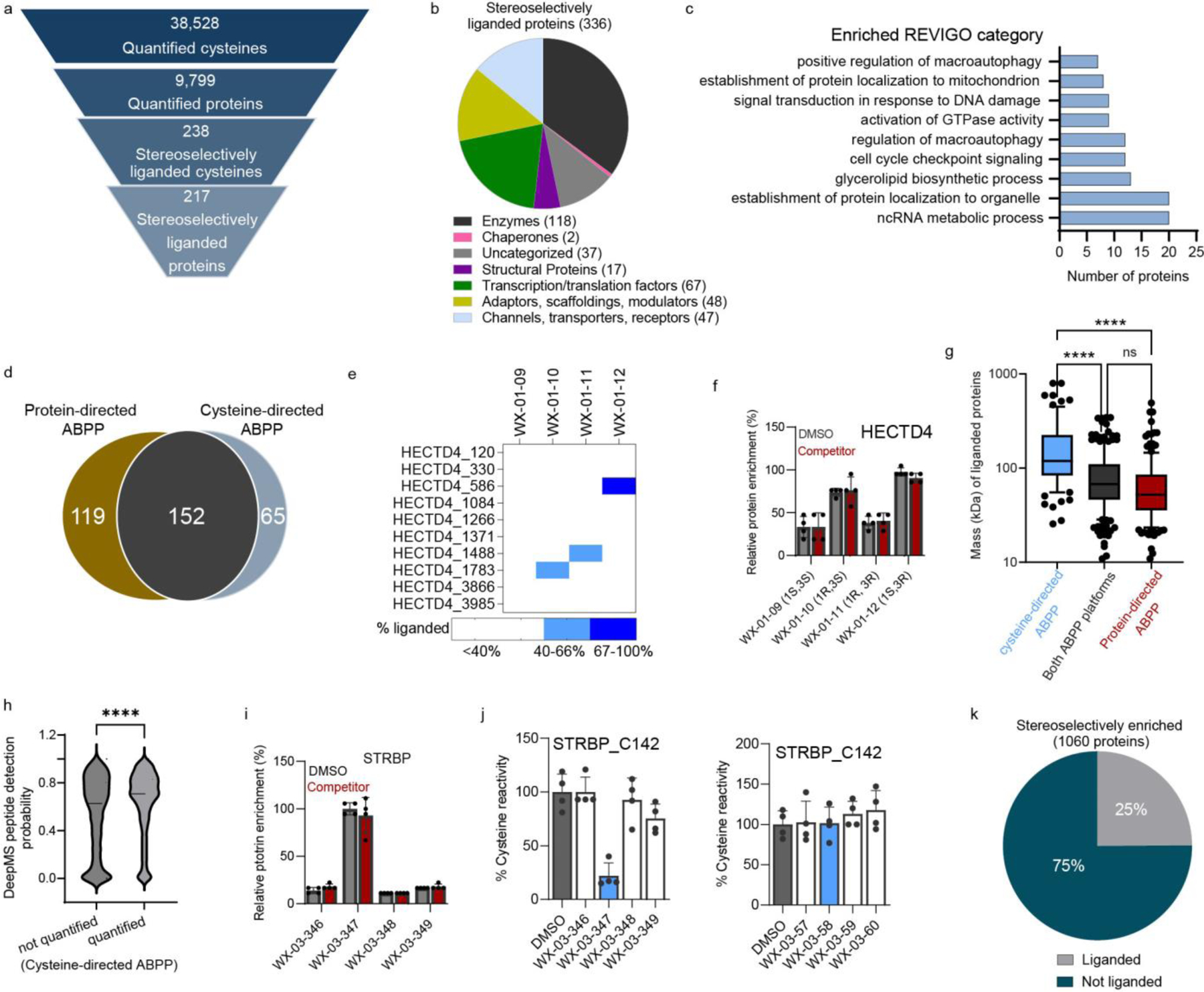
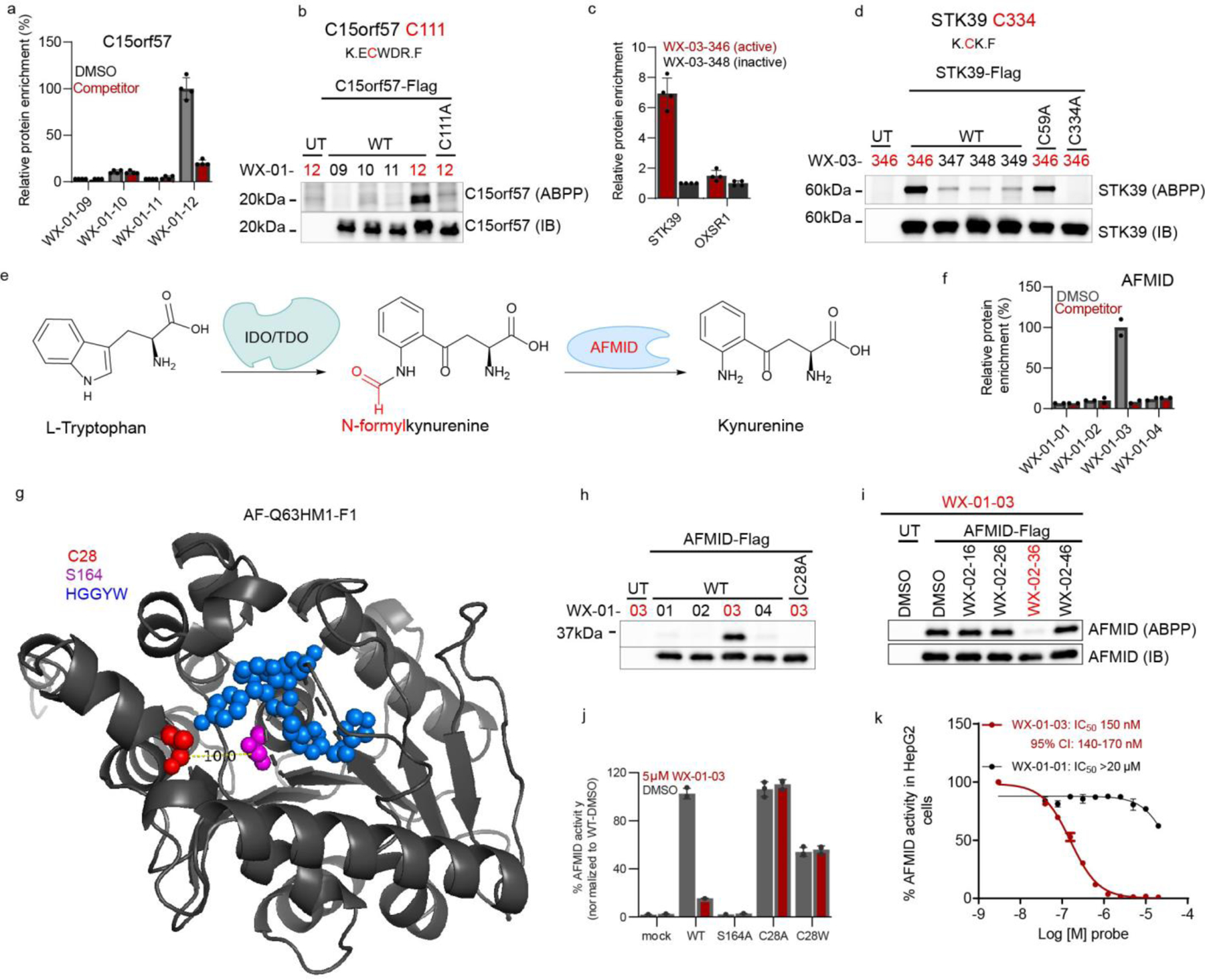
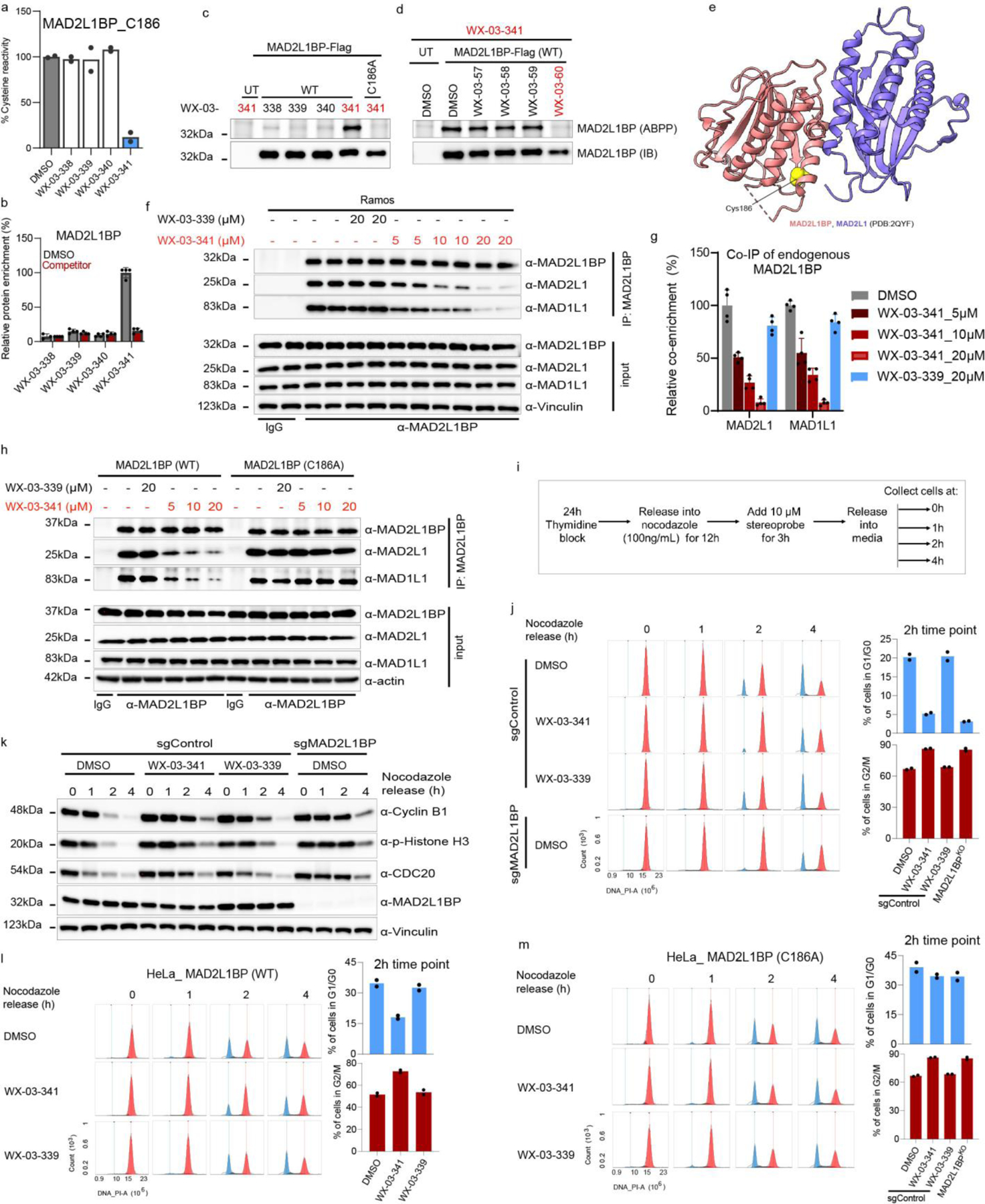
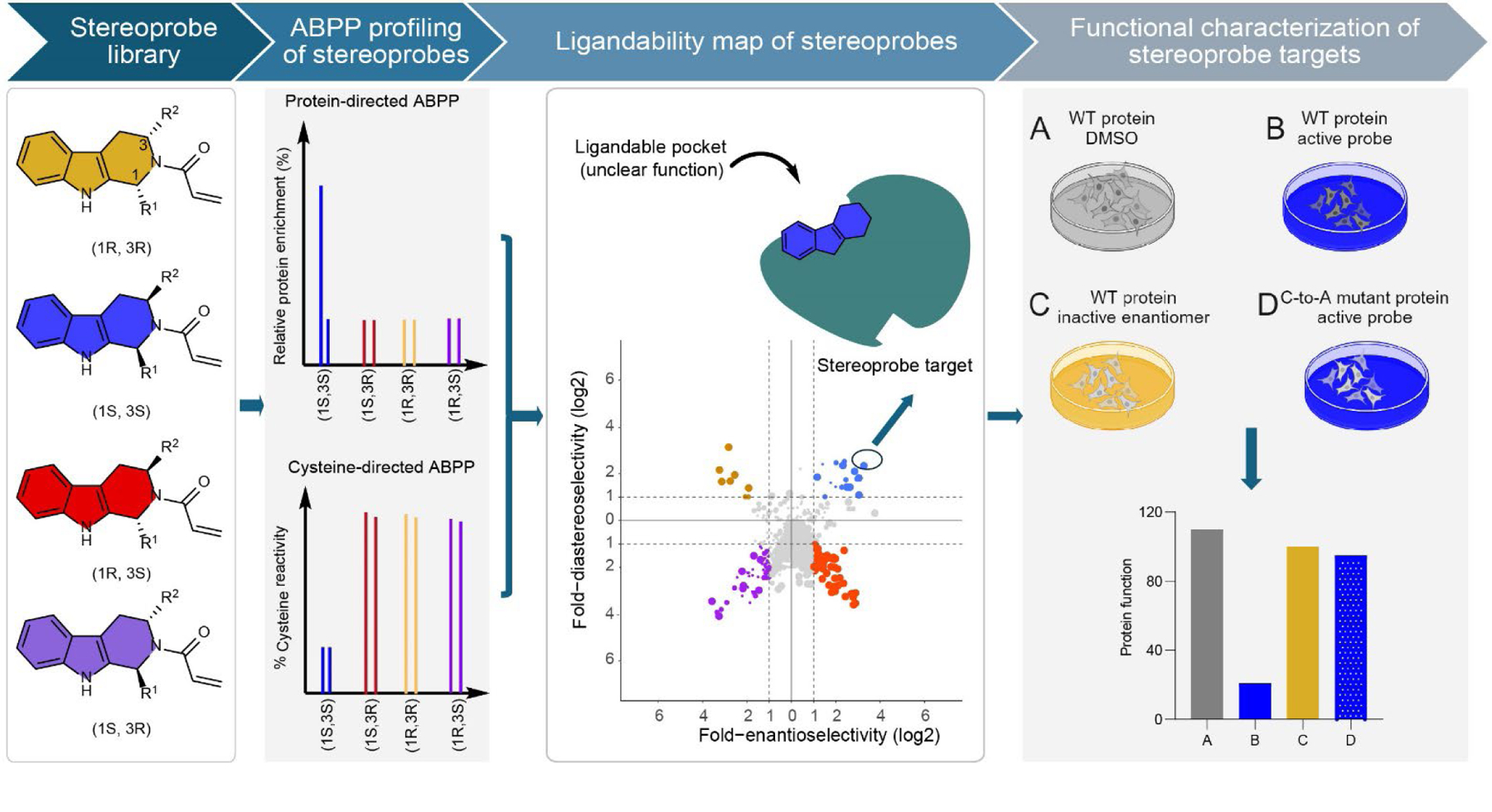
Covalent chemistry is a versatile approach for expanding the ligandability of the human proteome. Activity-based protein profiling (ABPP) can infer the specific residues modified by electrophilic compounds through competition with broadly reactive probes. However, the extent to which such residue-directed platforms fully assess the protein targets of electrophilic compounds in cells remains unclear. Here we evaluate a complementary protein-directed ABPP method that identifies proteins showing stereoselective reactivity with alkynylated, chiral electrophilic compounds-termed stereoprobes. Integration of protein- and cysteine-directed data from cancer cells treated with tryptoline acrylamide stereoprobes revealed generally well-correlated ligandability maps and highlighted features, such as protein size and the proteotypicity of cysteine-containing peptides, that explain gaps in each ABPP platform. In total, we identified stereoprobe binding events for >300 structurally and functionally diverse proteins, including compounds that stereoselectively and site-specifically disrupt MAD2L1BP interactions with the spindle assembly checkpoint complex leading to delayed mitotic exit in cancer cells.
© 2024. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing Interests
G.M.S., T. N., and P.A. are employees of Vividion Therapeutics, and B.F.C. is a founder and member of the Board of Directors of Vividion Therapeutics. The remaining authors declare no competing interests.
Figures

References
-
- Chakravarty D; Solit DB, Clinical cancer genomic profiling. Nat Rev Genet 2021, 22 (8), 483–501. - PubMed
-
- Wang ZZ; Shi XX; Huang GY; Hao GF; Yang GF, Fragment-based drug discovery supports drugging 'undruggable' protein-protein interactions. Trends Biochem Sci 2023, 48 (6), 539–552. - PubMed
-
- Lundquist KP; Panchal V; Gotfredsen CH; Brenk R; Clausen MH, Fragment-Based Drug Discovery for RNA Targets. ChemMedChem 2021, 16 (17), 2588–2603. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
