

Multiple aspects of amyloid dynamics in vivo integrate to establish prion variant dominance in yeast
- PMID: 39139213
- PMCID: PMC11319303
- DOI: 10.3389/fnmol.2024.1439442
Multiple aspects of amyloid dynamics in vivo integrate to establish prion variant dominance in yeast
Abstract
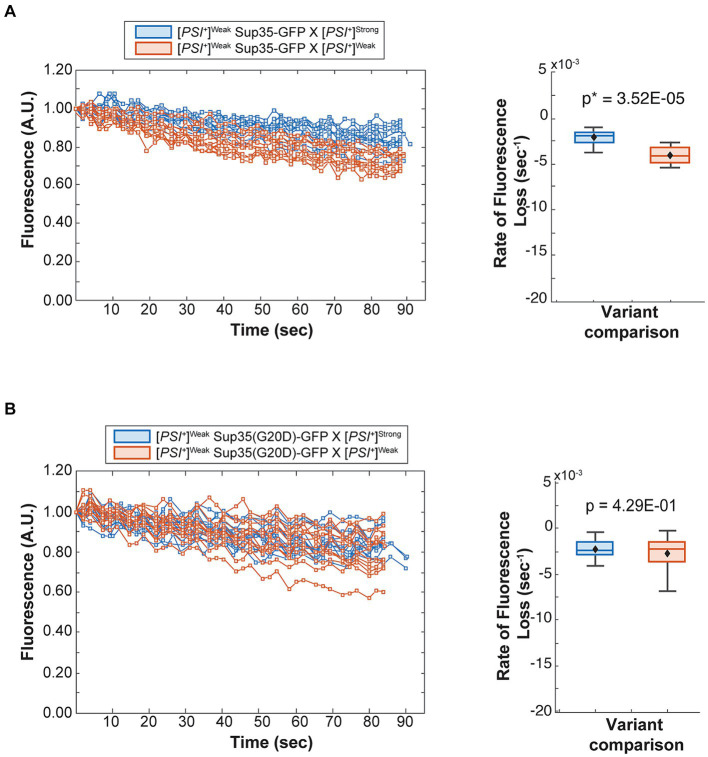
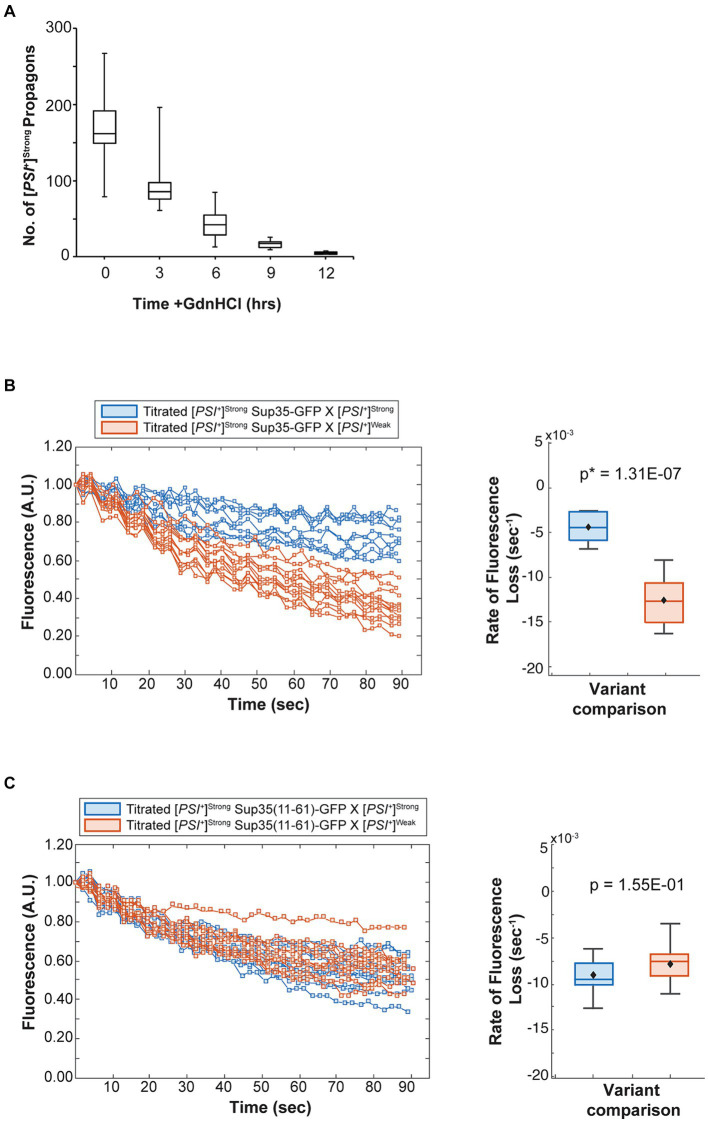
Prion variants are self-perpetuating conformers of a single protein that assemble into amyloid fibers and confer unique phenotypic states. Multiple prion variants can arise, particularly in response to changing environments, and interact within an organism. These interactions are often competitive, with one variant establishing phenotypic dominance over the others. This dominance has been linked to the competition for non-prion state protein, which must be converted to the prion state via a nucleated polymerization mechanism. However, the intrinsic rates of conversion, determined by the conformation of the variant, cannot explain prion variant dominance, suggesting a more complex interaction. Using the yeast prion system [PSI+ ], we have determined the mechanism of dominance of the [PSI+ ]Strong variant over the [PSI+ ]Weak variant in vivo. When mixed by mating, phenotypic dominance is established in zygotes, but the two variants persist and co-exist in the lineage descended from this cell. [PSI+ ]Strong propagons, the heritable unit, are amplified at the expense of [PSI+ ]Weak propagons, through the efficient conversion of soluble Sup35 protein, as revealed by fluorescence photobleaching experiments employing variant-specific mutants of Sup35. This competition, however, is highly sensitive to the fragmentation of [PSI+ ]Strong amyloid fibers, with even transient inhibition of the fragmentation catalyst Hsp104 promoting amplification of [PSI+ ]Weak propagons. Reducing the number of [PSI+ ]Strong propagons prior to mating, similarly promotes [PSI+ ]Weak amplification and conversion of soluble Sup35, indicating that template number and conversion efficiency combine to determine dominance. Thus, prion variant dominance is not an absolute hierarchy but rather an outcome arising from the dynamic interplay between unique protein conformations and their interactions with distinct cellular proteostatic niches.
Keywords: Hsp104; SUP35; amyloid; chaperone; prion; variant competition.
Copyright © 2024 Norton, Seah, Santiago, Sindi and Serio.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures




Similar articles
-
The NatA acetyltransferase couples Sup35 prion complexes to the [PSI+] phenotype.Mol Biol Cell. 2009 Feb;20(3):1068-80. doi: 10.1091/mbc.e08-04-0436. Epub 2008 Dec 10. Mol Biol Cell. 2009. PMID: 19073888 Free PMC article.
-
Heat shock protein 104 (Hsp104)-mediated curing of [PSI+] yeast prions depends on both [PSI+] conformation and the properties of the Hsp104 homologs.J Biol Chem. 2017 May 26;292(21):8630-8641. doi: 10.1074/jbc.M116.770719. Epub 2017 Apr 3. J Biol Chem. 2017. PMID: 28373280 Free PMC article.
-
The small heat shock protein Hsp31 cooperates with Hsp104 to modulate Sup35 prion aggregation.Prion. 2016 Nov;10(6):444-465. doi: 10.1080/19336896.2016.1234574. Prion. 2016. PMID: 27690738 Free PMC article.
-
Prion and nonprion amyloids: a comparison inspired by the yeast Sup35 protein.Prion. 2007 Jul-Sep;1(3):179-84. doi: 10.4161/pri.1.3.4840. Epub 2007 Jul 6. Prion. 2007. PMID: 19164899 Free PMC article. Review.
-
Self-perpetuating changes in Sup35 protein conformation as a mechanism of heredity in yeast.Biochem Soc Symp. 2001;(68):35-43. doi: 10.1042/bss0680035. Biochem Soc Symp. 2001. PMID: 11573346 Review.
References
-
- Bartz J. C., Bessen R. A., McKenzie D., Marsh R. F., Aiken J. M. (2000). Adaptation and selection of prion protein strain conformations following interspecies transmission of transmissible mink encephalopathy. J. Virol. 74, 5542–5547. doi: 10.1128/JVI.74.12.5542-5547.2000, PMID: - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources