

The genomes of all lungfish inform on genome expansion and tetrapod evolution
- PMID: 39143221
- PMCID: PMC11514621
- DOI: 10.1038/s41586-024-07830-1
The genomes of all lungfish inform on genome expansion and tetrapod evolution
Abstract
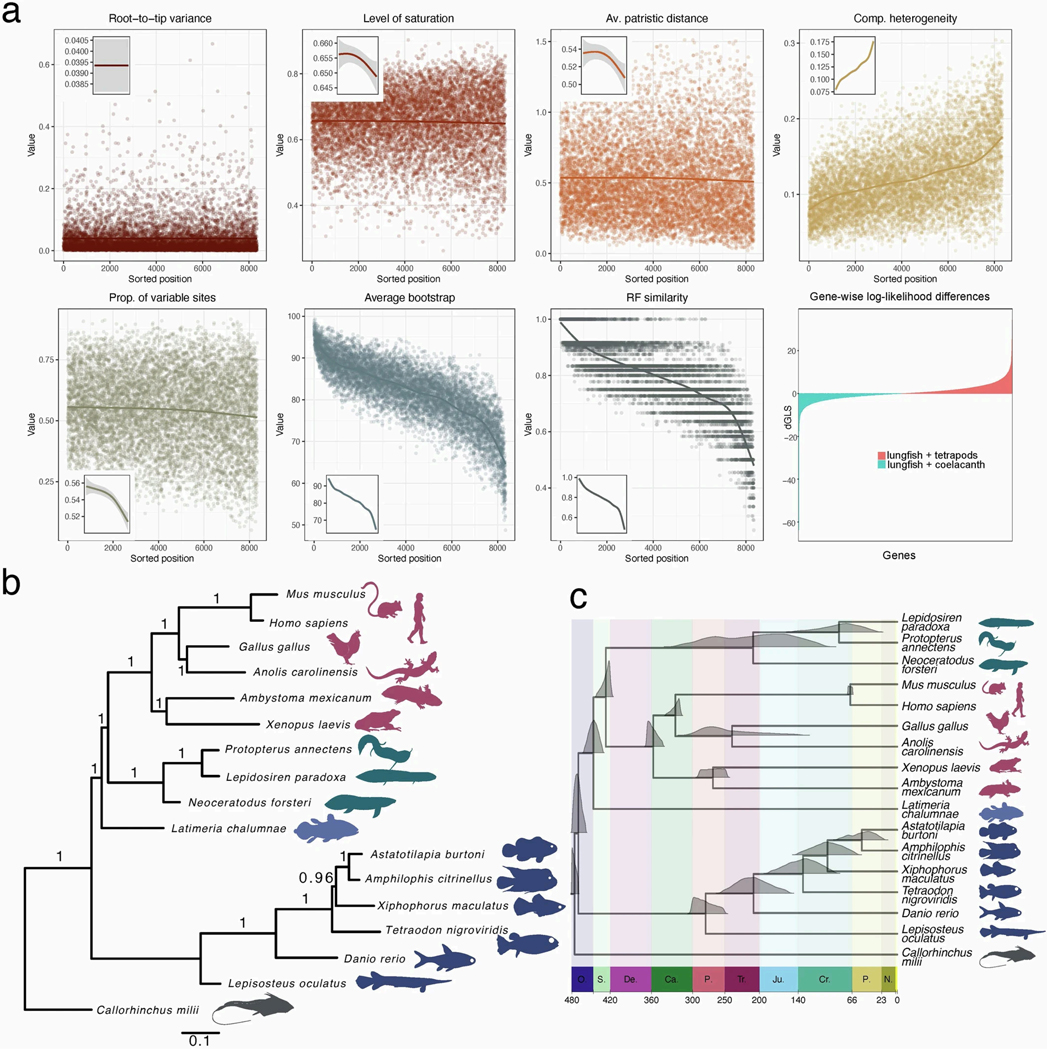
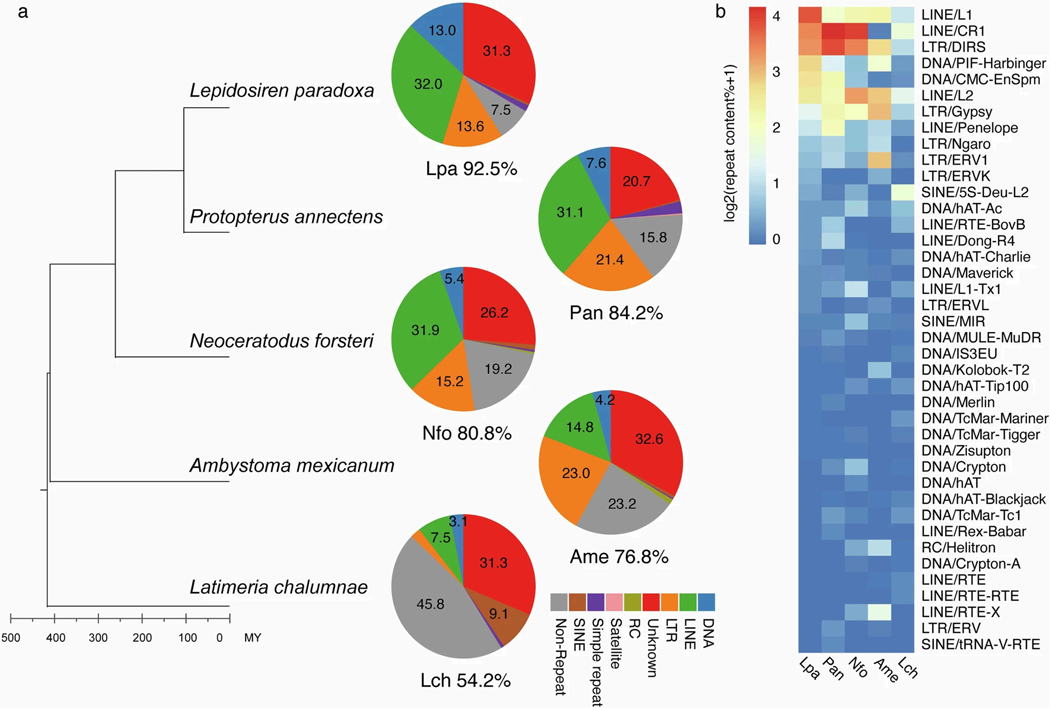
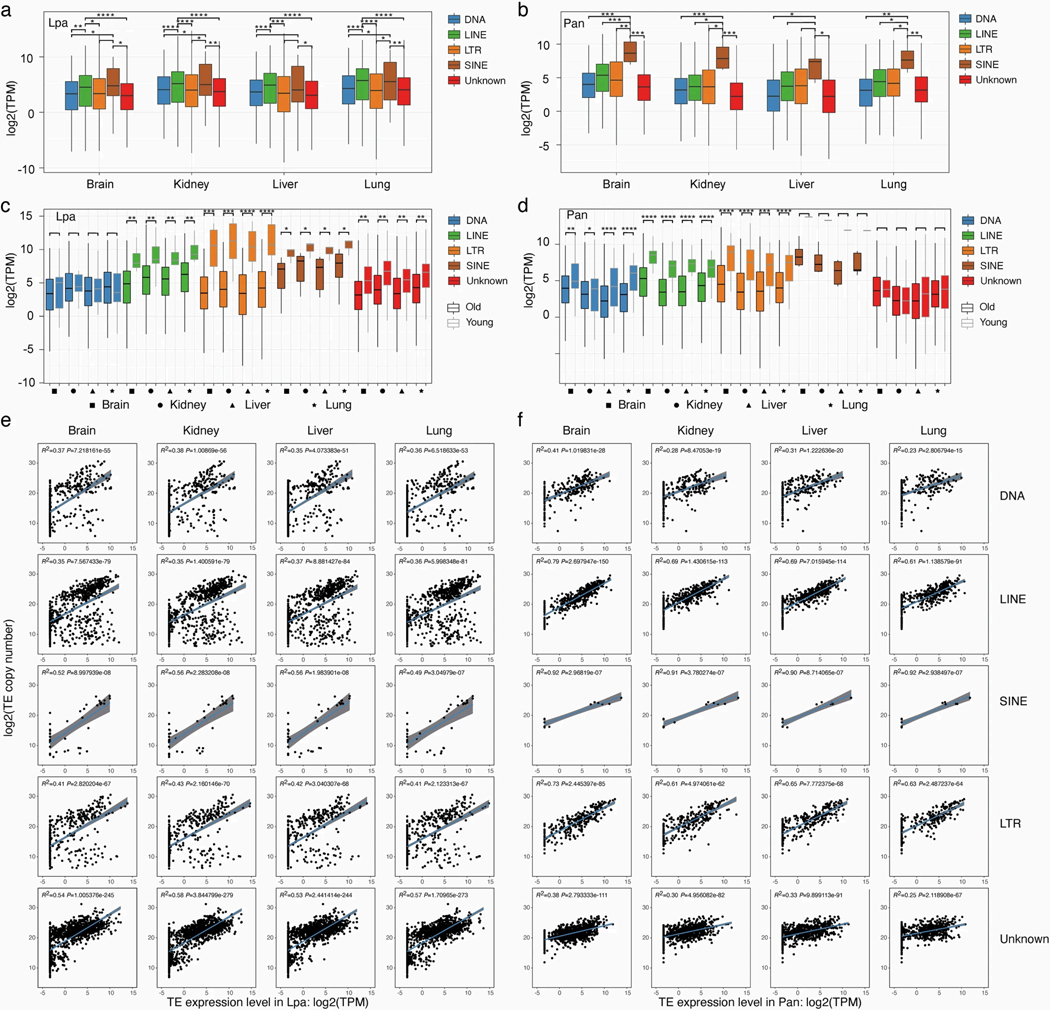
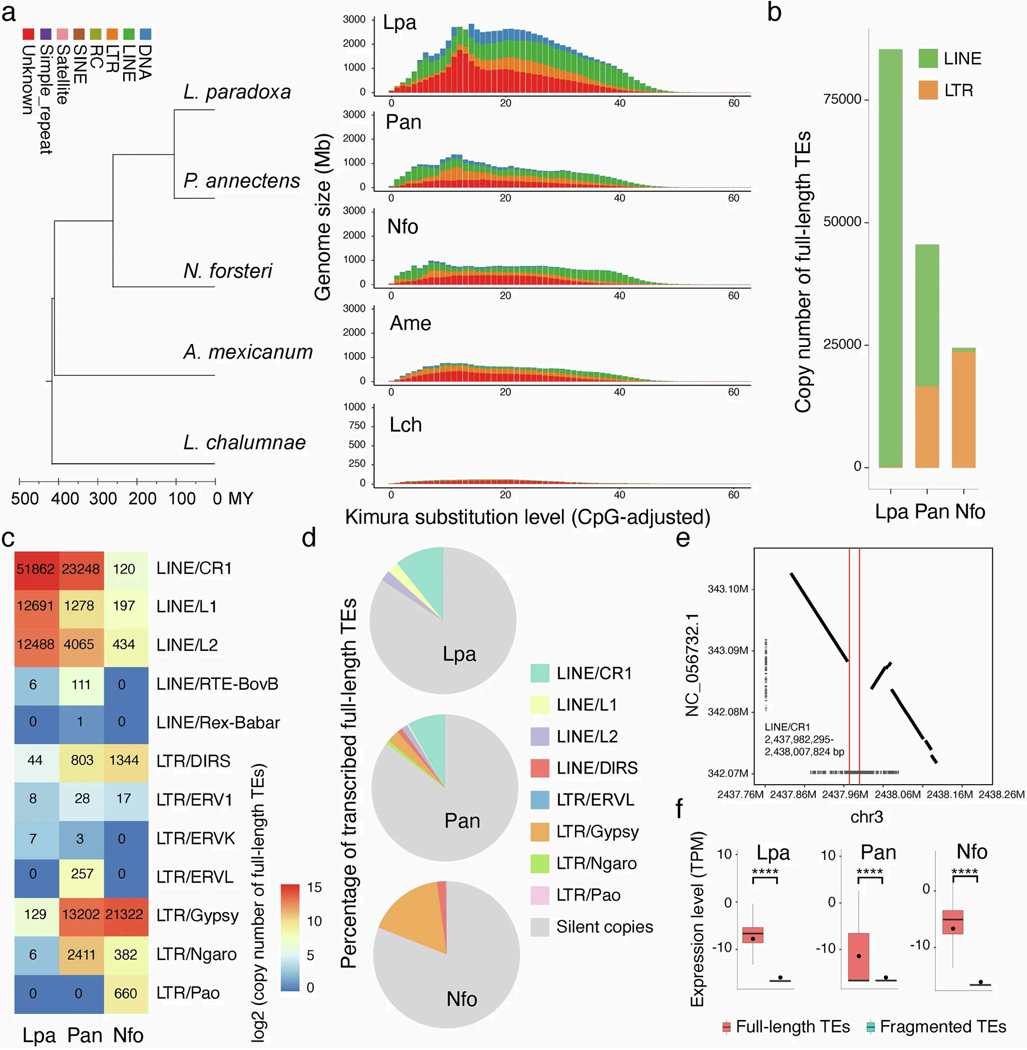
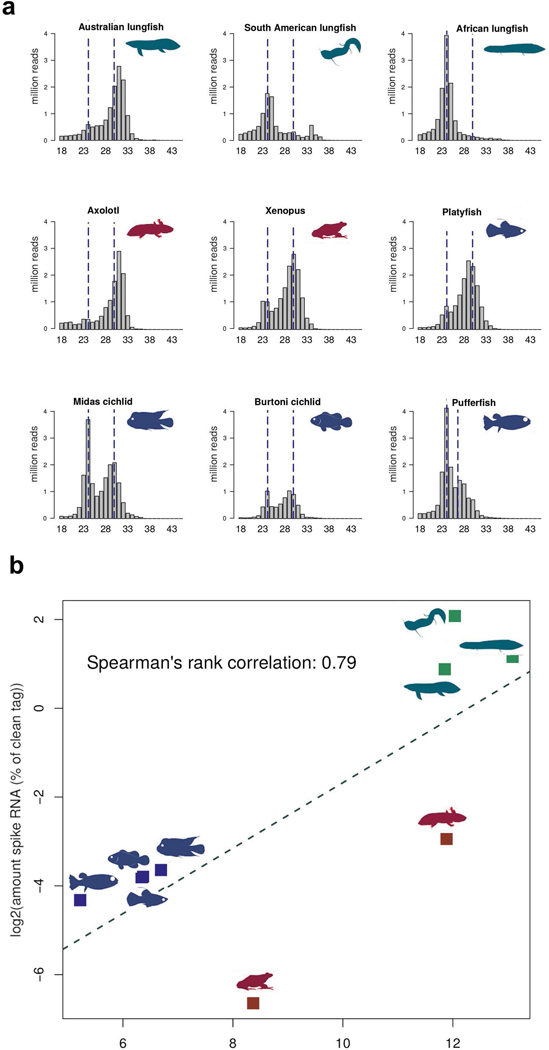
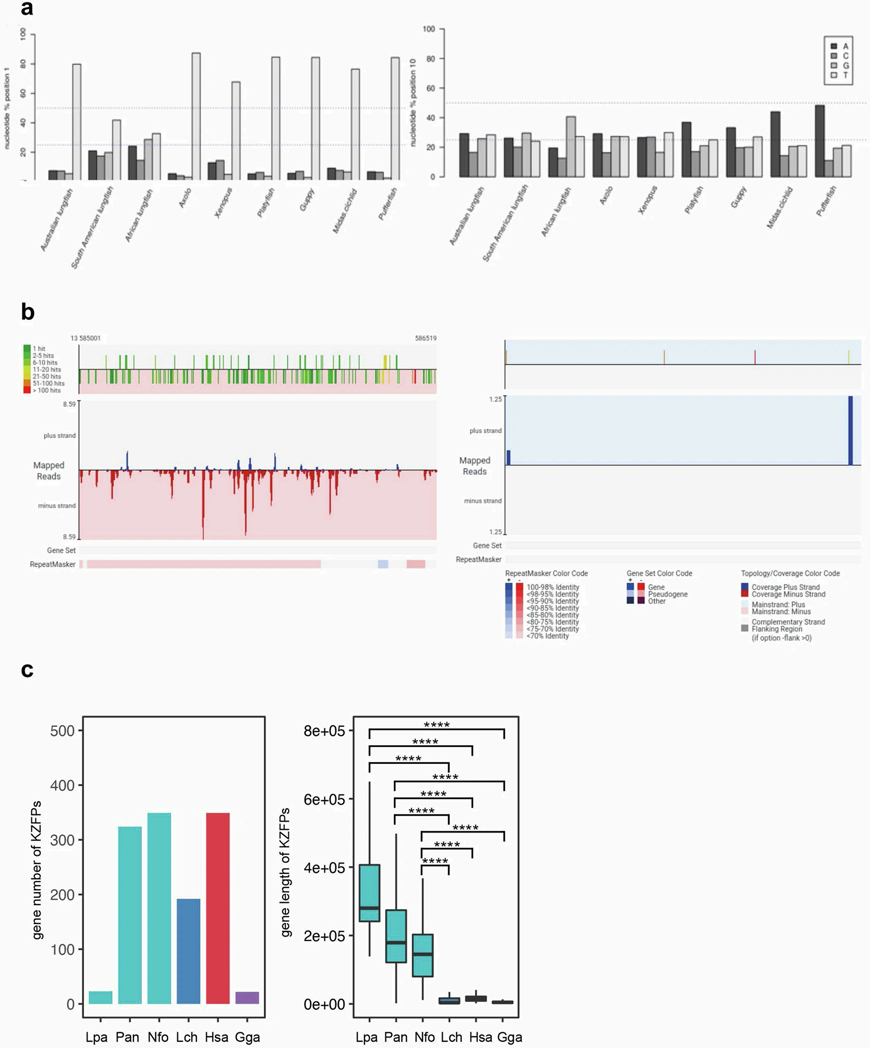
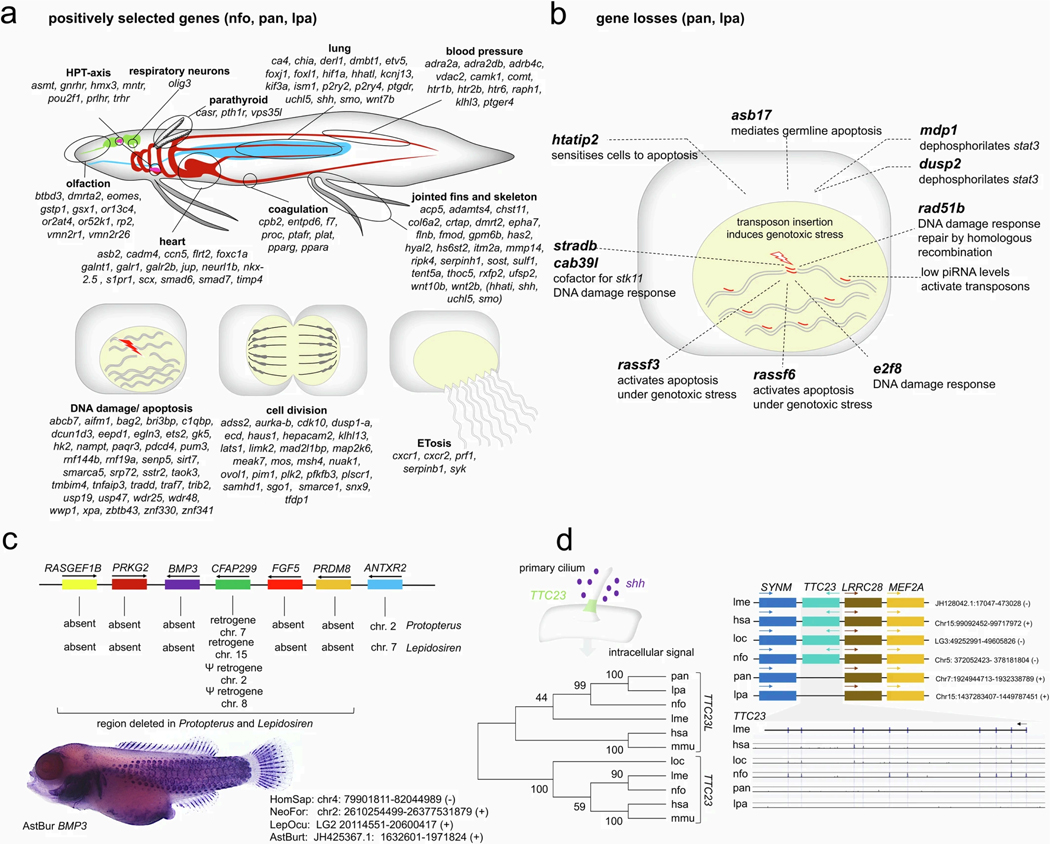
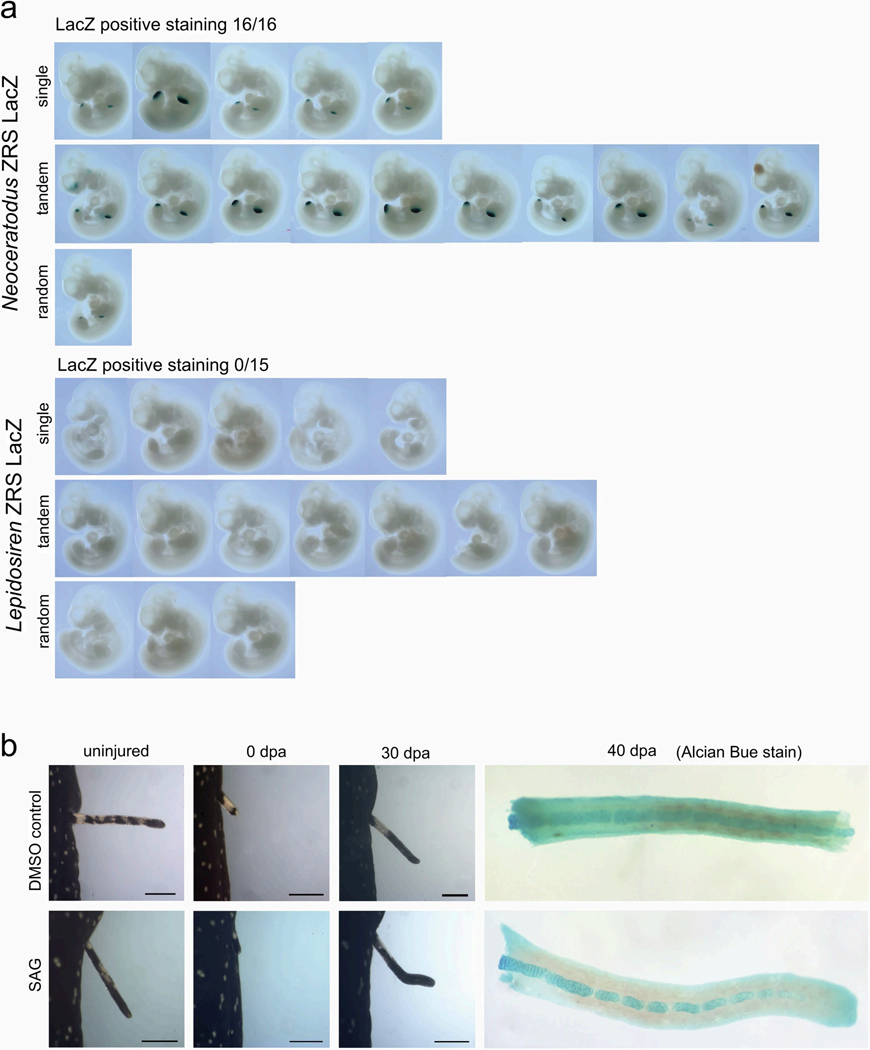
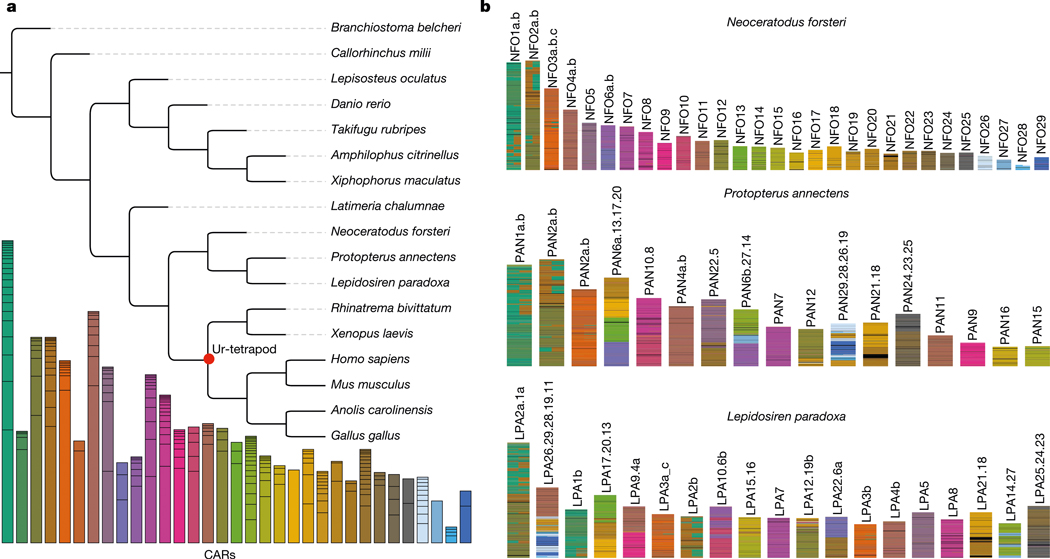
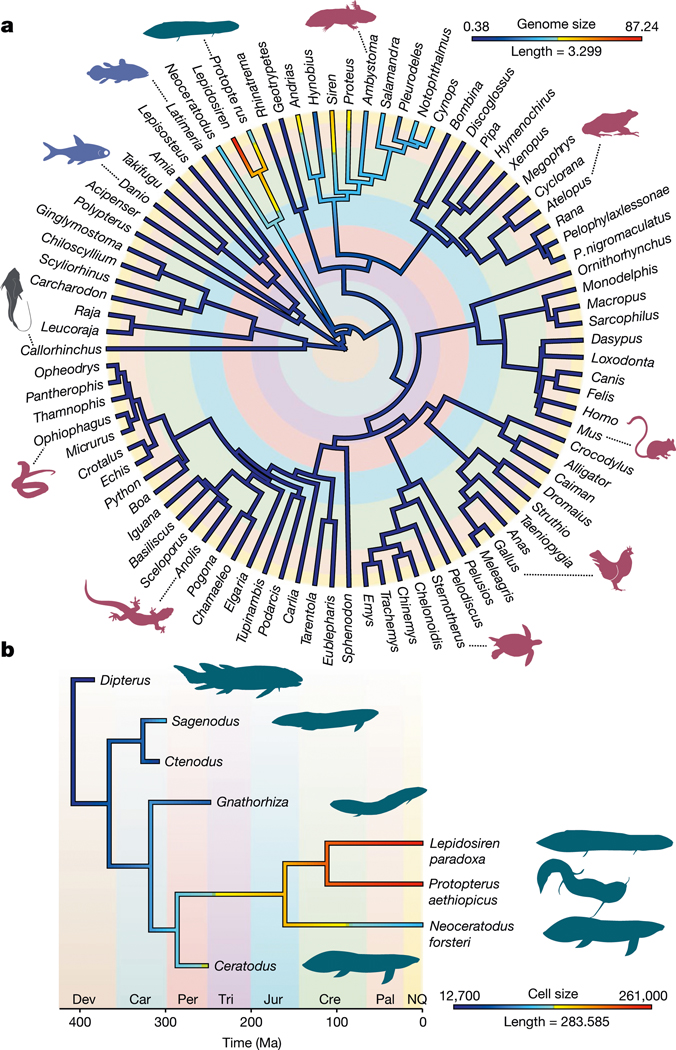
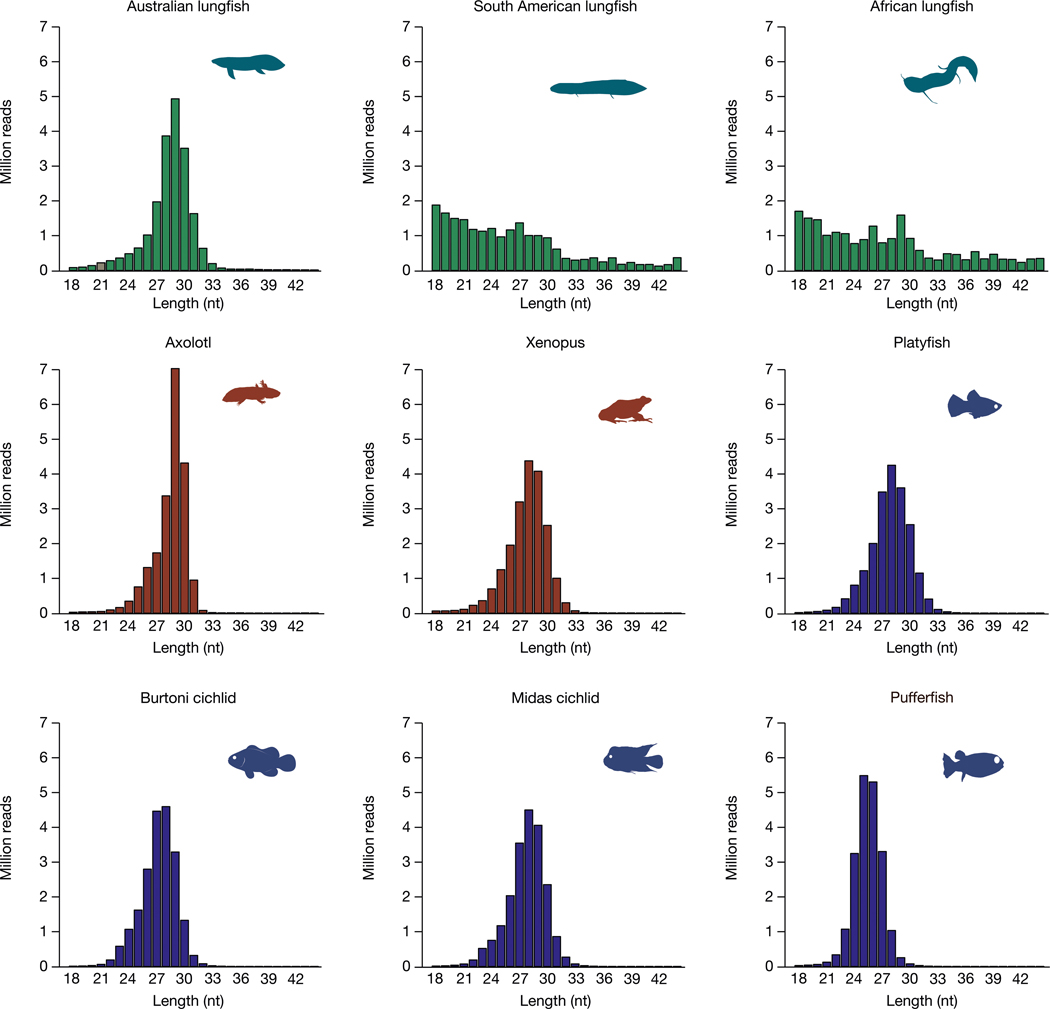
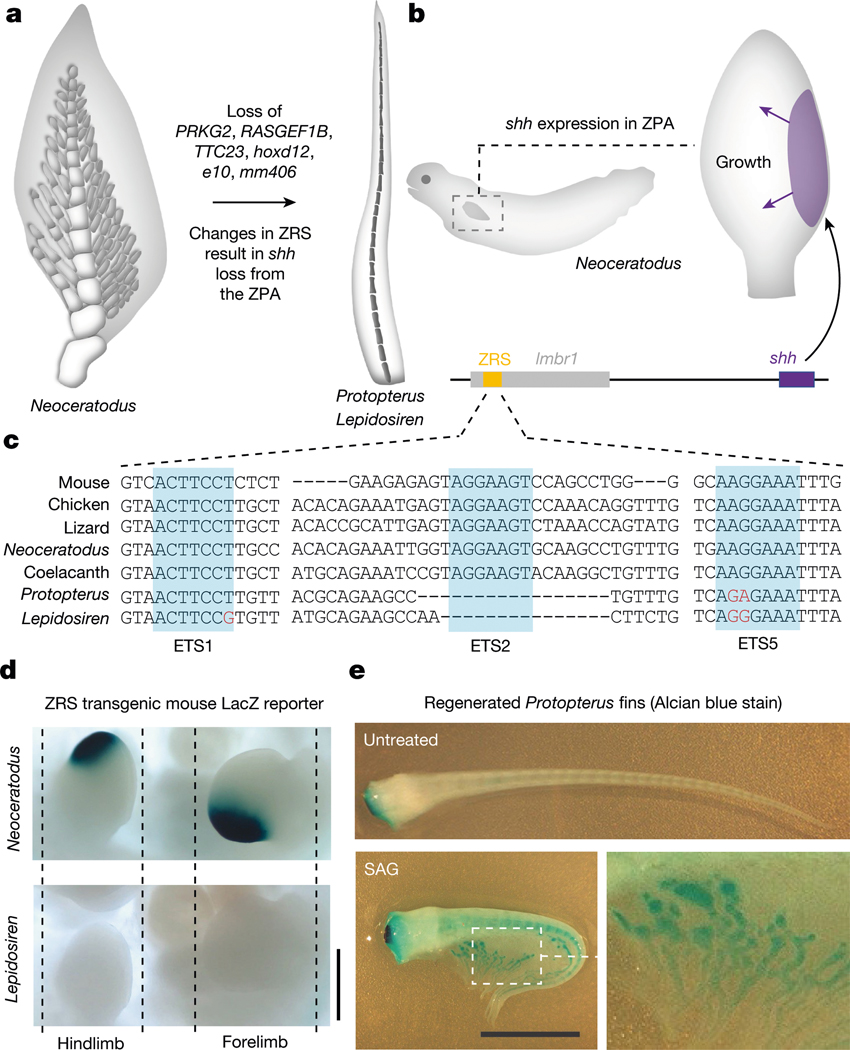
The genomes of living lungfishes can inform on the molecular-developmental basis of the Devonian sarcopterygian fish-tetrapod transition. We de novo sequenced the genomes of the African (Protopterus annectens) and South American lungfishes (Lepidosiren paradoxa). The Lepidosiren genome (about 91 Gb, roughly 30 times the human genome) is the largest animal genome sequenced so far and more than twice the size of the Australian (Neoceratodus forsteri)1 and African2 lungfishes owing to enlarged intergenic regions and introns with high repeat content (about 90%). All lungfish genomes continue to expand as some transposable elements (TEs) are still active today. In particular, Lepidosiren's genome grew extremely fast during the past 100 million years (Myr), adding the equivalent of one human genome every 10 Myr. This massive genome expansion seems to be related to a reduction of PIWI-interacting RNAs and C2H2 zinc-finger and Krüppel-associated box (KRAB)-domain protein genes that suppress TE expansions. Although TE abundance facilitates chromosomal rearrangements, lungfish chromosomes still conservatively reflect the ur-tetrapod karyotype. Neoceratodus' limb-like fins still resemble those of their extinct relatives and remained phenotypically static for about 100 Myr. We show that the secondary loss of limb-like appendages in the Lepidosiren-Protopterus ancestor was probably due to loss of sonic hedgehog limb-specific enhancers.
© 2024. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Figures


References
-
- Wang K et al. African lungfish genome sheds light on the vertebrate water-to-land transition. Cell 184, 1362–1376.e1318 (2021). - PubMed
-
- Krefft JLG Description of a gigantic amphibian allied to the genus Lepidosiren from the Wide-Bay district, Queensland. Proc. Zool. Soc. Lond 1870, 221–224 (1870).
-
- Meyer A & Dolven SI Molecules, fossils, and the origin of tetrapods. J. Mol. Evol 35, 102–113 (1992). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
