

Olfactory sensory neuron population expansions influence projection neuron adaptation and enhance odour tracking
- PMID: 39147786
- PMCID: PMC11327376
- DOI: 10.1038/s41467-024-50808-w
Olfactory sensory neuron population expansions influence projection neuron adaptation and enhance odour tracking
Abstract
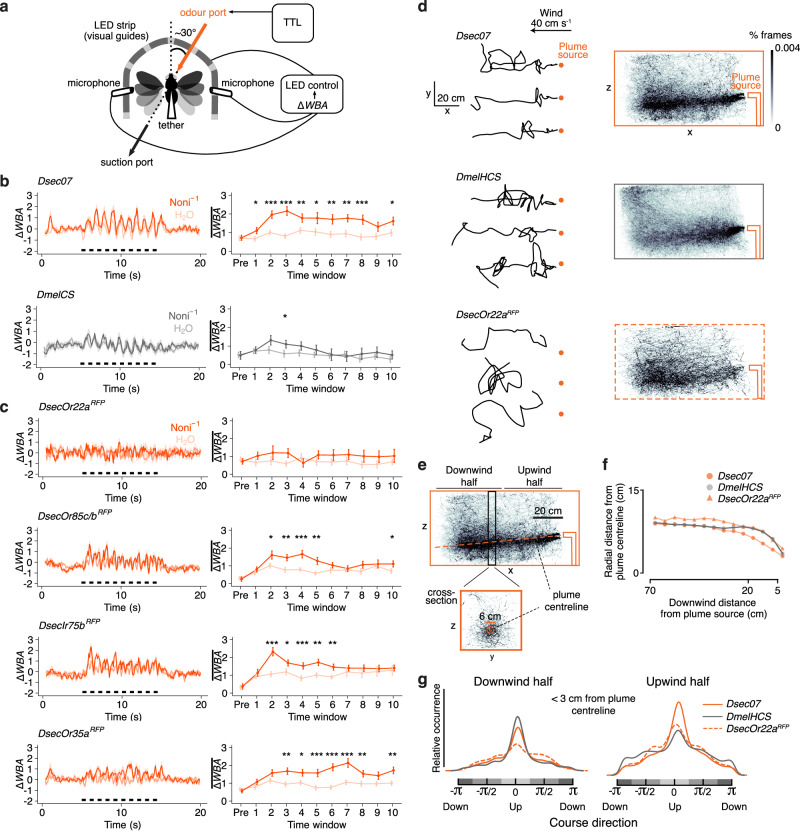
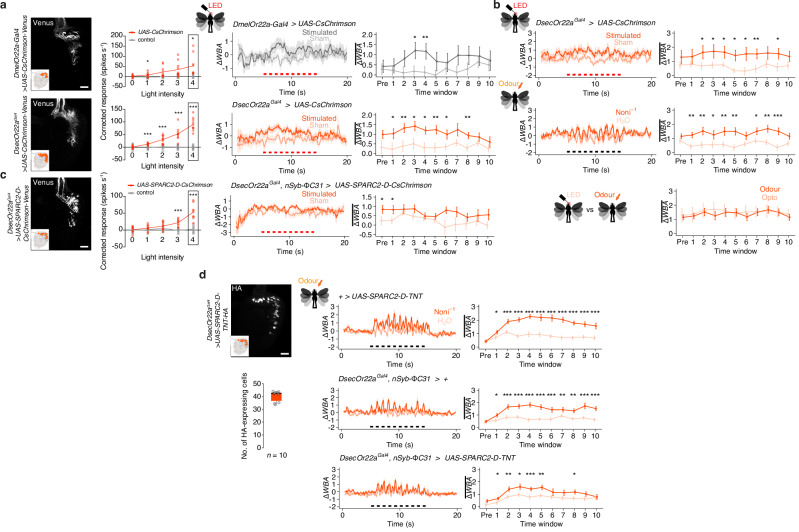
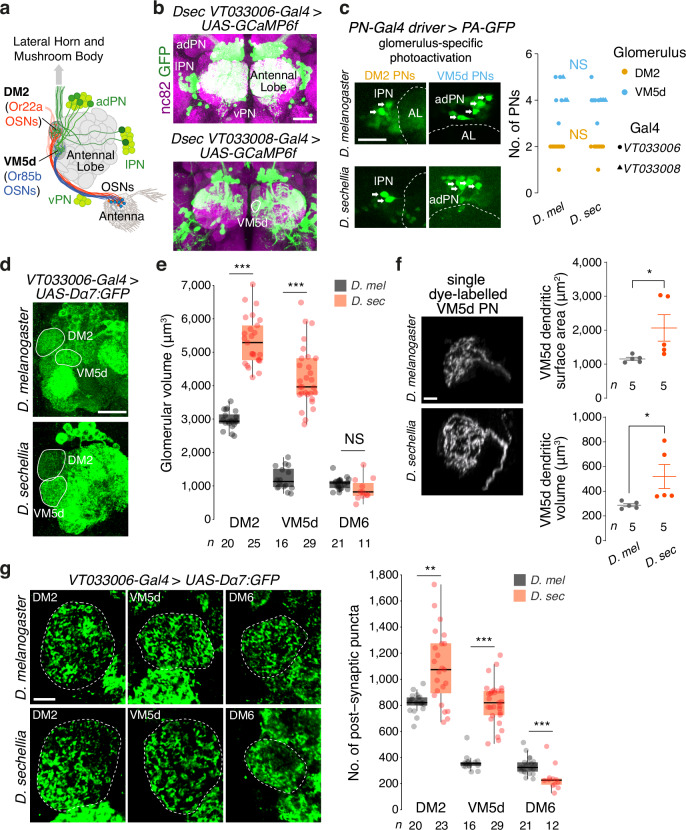
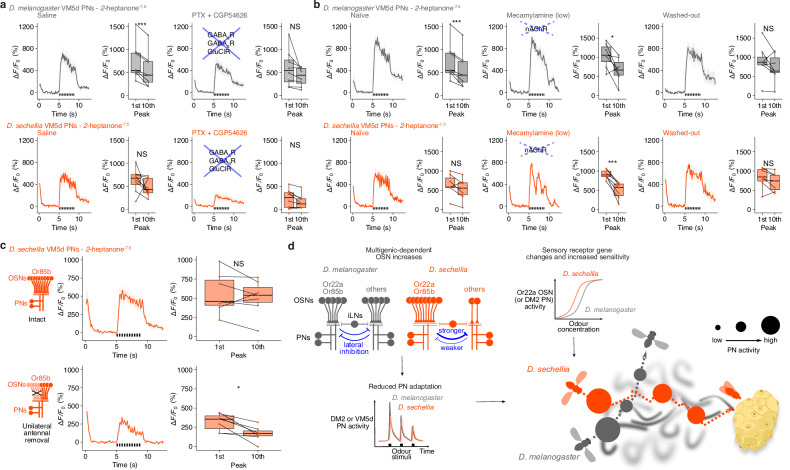
The evolutionary expansion of sensory neuron populations detecting important environmental cues is widespread, but functionally enigmatic. We investigated this phenomenon through comparison of homologous olfactory pathways of Drosophila melanogaster and its close relative Drosophila sechellia, an extreme specialist for Morinda citrifolia noni fruit. D. sechellia has evolved species-specific expansions in select, noni-detecting olfactory sensory neuron (OSN) populations, through multigenic changes. Activation and inhibition of defined proportions of neurons demonstrate that OSN number increases contribute to stronger, more persistent, noni-odour tracking behaviour. These expansions result in increased synaptic connections of sensory neurons with their projection neuron (PN) partners, which are conserved in number between species. Surprisingly, having more OSNs does not lead to greater odour-evoked PN sensitivity or reliability. Rather, pathways with increased sensory pooling exhibit reduced PN adaptation, likely through weakened lateral inhibition. Our work reveals an unexpected functional impact of sensory neuron population expansions to explain ecologically-relevant, species-specific behaviour.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Update of
-
Sensory neuron population expansion enhances odor tracking without sensitizing projection neurons.bioRxiv [Preprint]. 2024 Feb 28:2023.09.15.556782. doi: 10.1101/2023.09.15.556782. bioRxiv. 2024. Update in: Nat Commun. 2024 Aug 15;15(1):7041. doi: 10.1038/s41467-024-50808-w. PMID: 37745467 Free PMC article. Updated. Preprint.
References
MeSH terms
Grants and funding
- 310030B-185377/Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (Swiss National Science Foundation)
- ALTF 454-2019/European Molecular Biology Organization (EMBO)
- 202360258/MEXT | Japan Society for the Promotion of Science (JSPS)
- PZ00P3 185743/Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (Swiss National Science Foundation)
- P40 OD018537/OD/NIH HHS/United States
- 310030_201188/Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (Swiss National Science Foundation)
- R01 NS116584/NS/NINDS NIH HHS/United States
- PP00P3_176956/Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (Swiss National Science Foundation)
- LT000461/2015-L/Human Frontier Science Program (HFSP)
- 615094/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 European Research Council (H2020 Excellent Science - European Research Council)
- 833548/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 European Research Council (H2020 Excellent Science - European Research Council)
- FA9550-21-0122/United States Department of Defense | United States Air Force | AFMC | Air Force Office of Scientific Research (AF Office of Scientific Research)
- R01 DC018570/DC/NIDCD NIH HHS/United States
- R01EY022638/U.S. Department of Health & Human Services | National Institutes of Health (NIH)
- R01 EY022638/EY/NEI NIH HHS/United States
- 836783/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 Marie Skłodowska-Curie Actions (H2020 Excellent Science - Marie Skłodowska-Curie Actions)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
