

This is a preprint.
Persistent changes in nociceptor translatomes govern hyperalgesic priming in mouse models
- PMID: 39149295
- PMCID: PMC11326310
- DOI: 10.1101/2024.08.07.606891
Persistent changes in nociceptor translatomes govern hyperalgesic priming in mouse models
Update in
-
Persistent changes in the dorsal root ganglion nociceptor translatome governs hyperalgesic priming in mice: roles of GPR88 and Meteorin.Pain. 2025 Jun 1;166(6):1395-1405. doi: 10.1097/j.pain.0000000000003523. Epub 2025 Jan 28. Pain. 2025. PMID: 39878635
Abstract
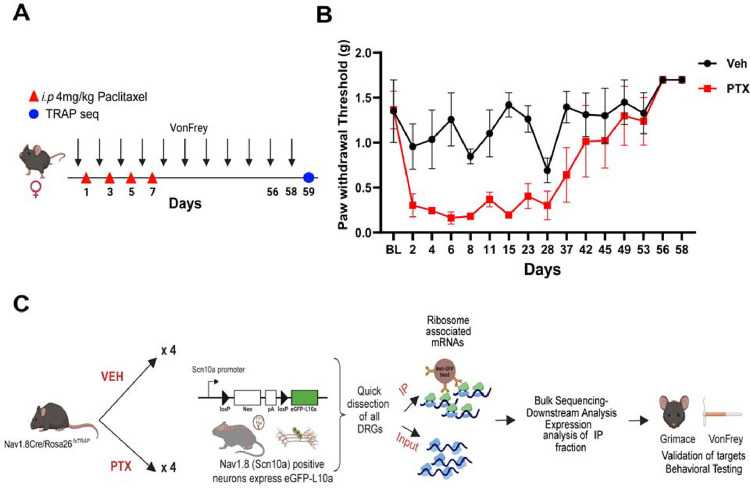
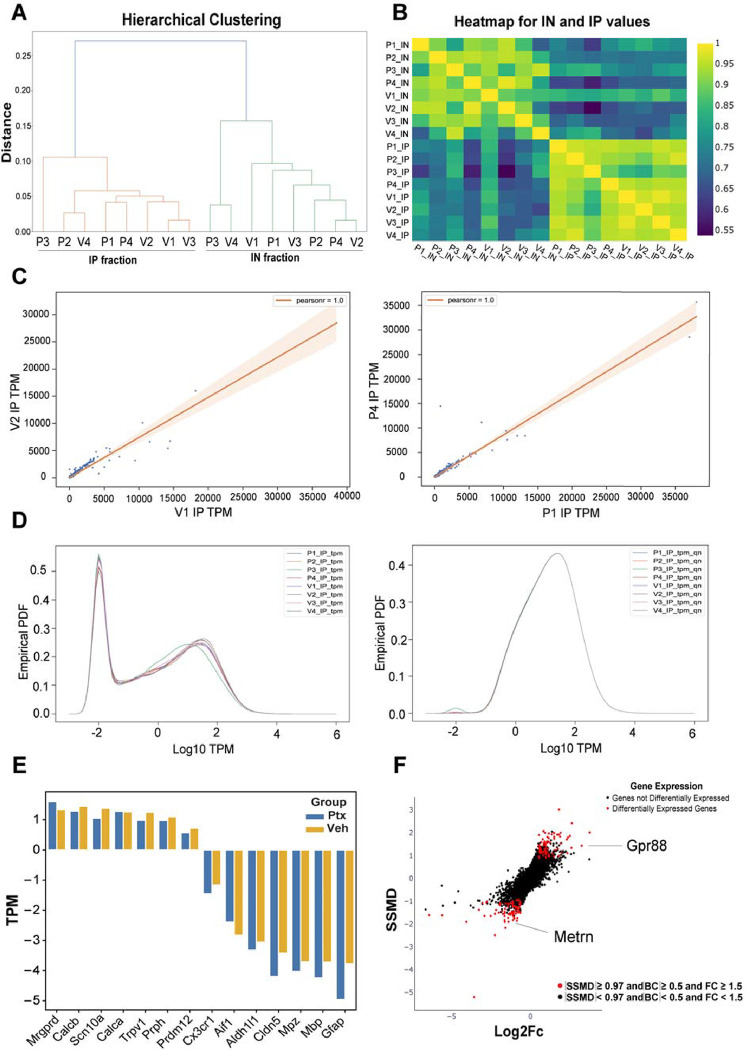
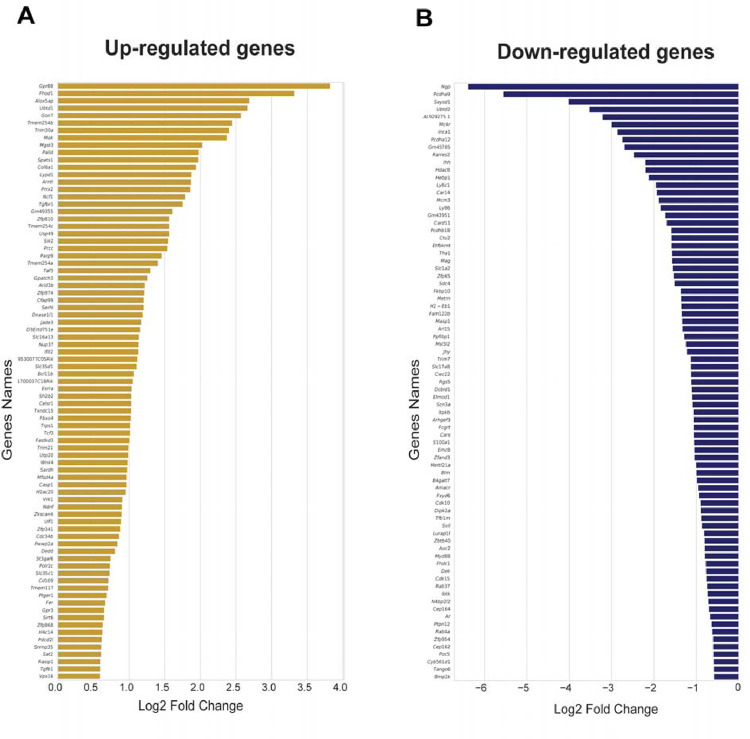
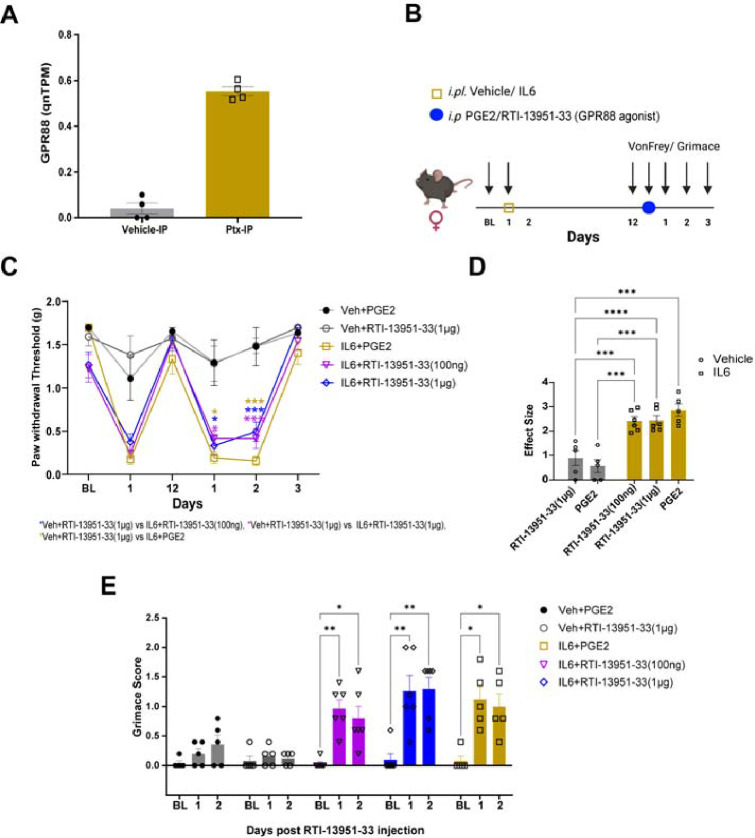
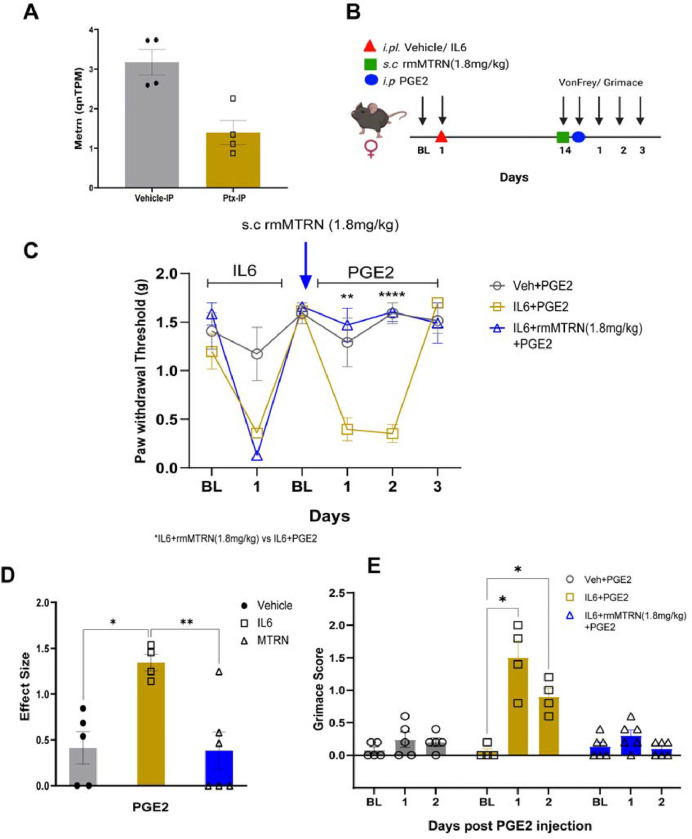
Hyperalgesic priming is a model system that has been widely used to understand plasticity in painful stimulus-detecting sensory neurons, called nociceptors. A key feature of this model system is that following priming, stimuli that do not normally cause hyperalgesia now readily provoke this state. We hypothesized that hyperalgesic priming occurs due to reorganization of translation of mRNA in nociceptors. To test this hypothesis, we used paclitaxel treatment as the priming stimulus and translating ribosome affinity purification (TRAP) to measure persistent changes in mRNA translation in Nav1.8+ nociceptors. TRAP sequencing revealed 161 genes with persistently altered mRNA translation in the primed state. We identified Gpr88 as upregulated and Metrn as downregulated. We confirmed a functional role for these genes, wherein a GPR88 agonist causes pain only in primed mice and established hyperalgesic priming is reversed by Meteorin. Our work demonstrates that altered nociceptor translatomes are causative in producing hyperalgesic priming.
Keywords: GPR88; IL6-mediated hyperalgesic priming; Meteorin; chemotherapy-induced peripheral neuropathy; hyperalgesic priming; translating ribosome affinity purification.
Conflict of interest statement
Conflict of Interest Statement: G Munro and KA Petersen are employees of Hoba Therapeutics. The authors declare no other conflicts of interest.
Figures
References
-
- Koltzenburg M., The changing sensitivity in the life of the nociceptor. Pain Suppl 6, S93–102 (1999); published online EpubAug ( - PubMed
-
- North R. Y., Li Y., Ray P., Rhines L. D., Tatsui C. E., Rao G., Johansson C. A., Zhang H., Kim Y. H., Zhang B., Dussor G., Kim T. H., Price T. J., Dougherty P. M., Electrophysiological and transcriptomic correlates of neuropathic pain in human dorsal root ganglion neurons. Brain 142, 1215–1226 (2019); published online EpubMay 1 ( 10.1093/brain/awz063). - DOI - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases