

This is a preprint.
A dorsal hippocampus-prodynorphinergic dorsolateral septum-to-lateral hypothalamus circuit mediates contextual gating of feeding
- PMID: 39149322
- PMCID: PMC11326193
- DOI: 10.1101/2024.08.02.606427
A dorsal hippocampus-prodynorphinergic dorsolateral septum-to-lateral hypothalamus circuit mediates contextual gating of feeding
Abstract
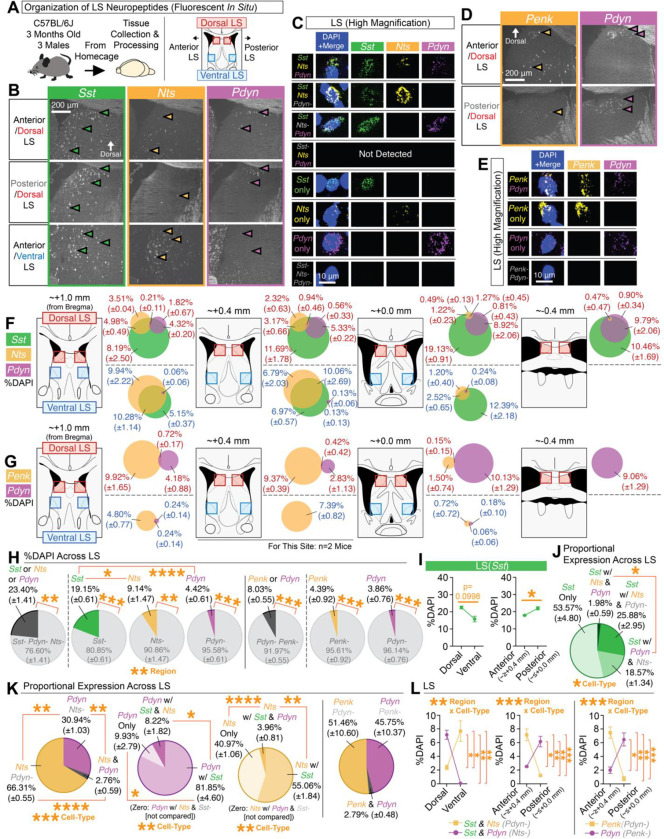
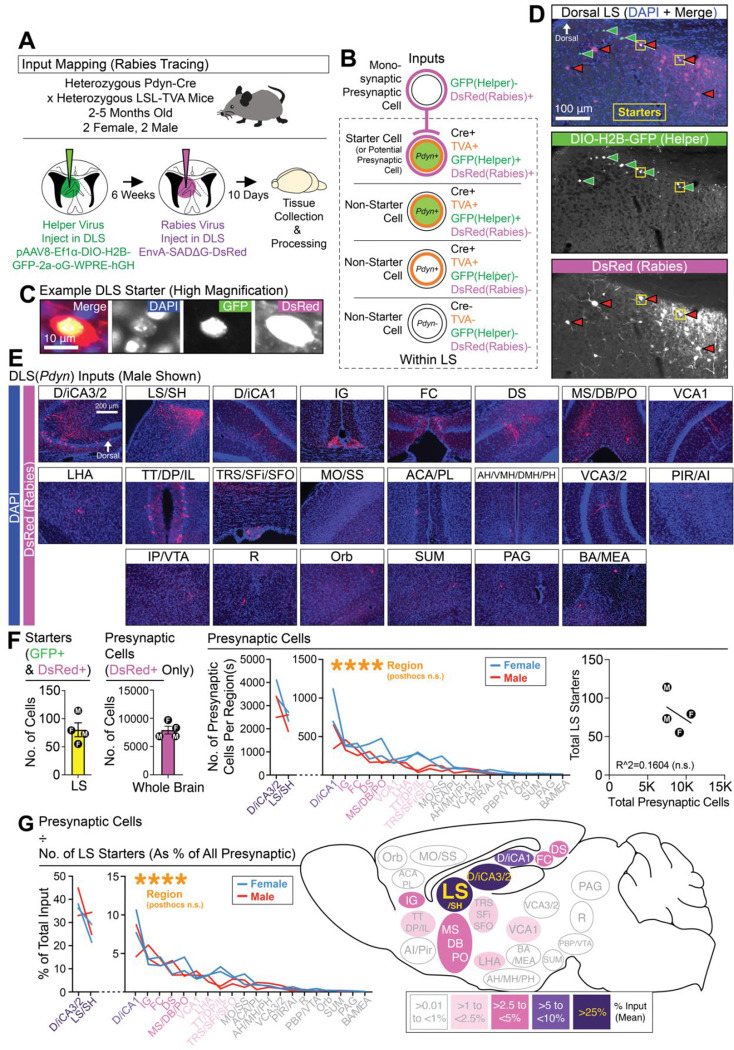
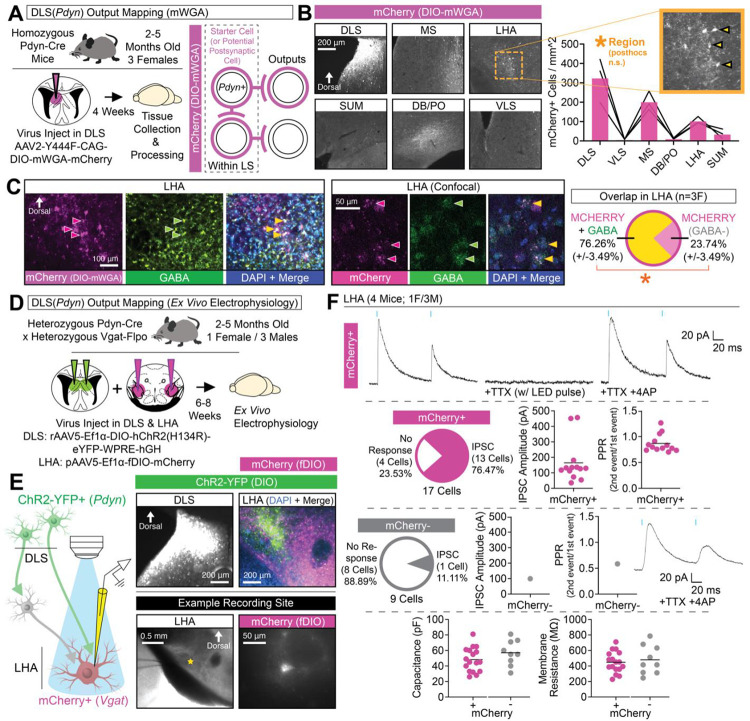
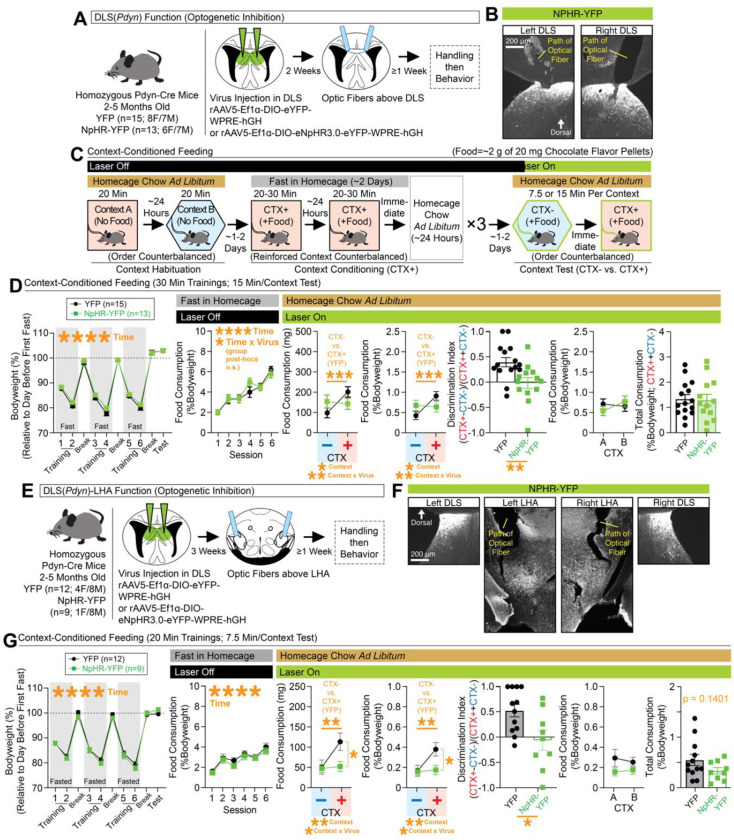
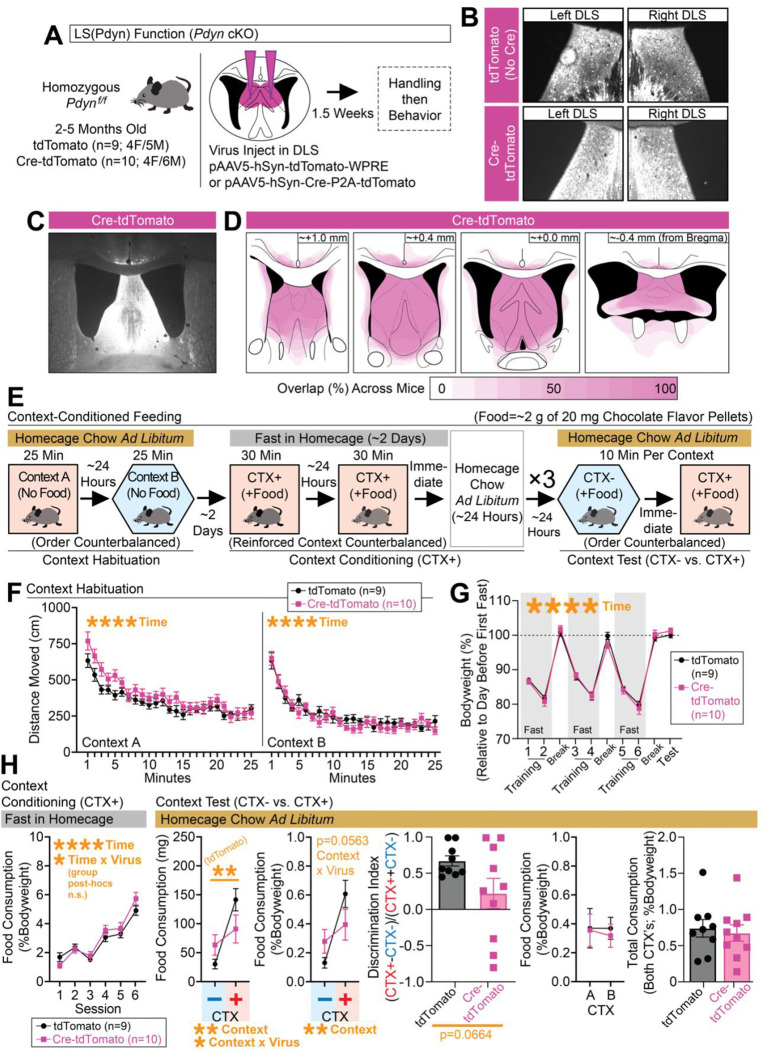
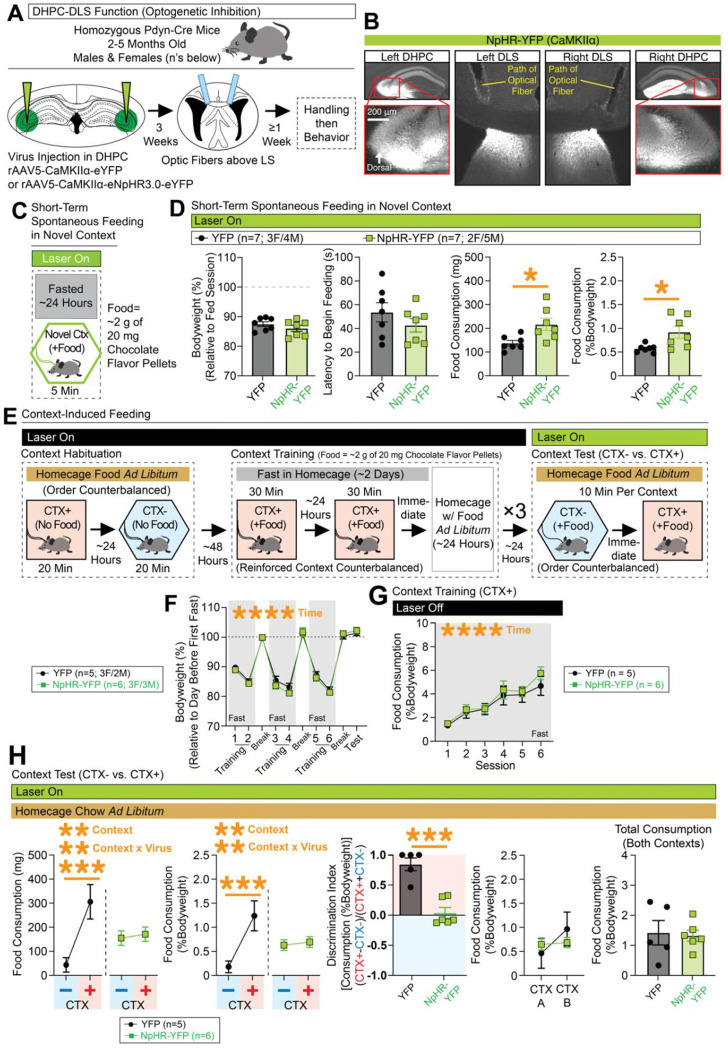
Adaptive regulation of feeding depends on linkage of internal states and food outcomes with contextual cues. Human brain imaging has identified dysregulation of a hippocampal-lateral hypothalamic area (LHA) network in binge eating, but mechanistic instantiation of underlying cell-types and circuitry is lacking. Here, we identify an evolutionary conserved and discrete Prodynorphin (Pdyn)-expressing subpopulation of Somatostatin (Sst)-expressing inhibitory neurons in the dorsolateral septum (DLS) that receives primarily dorsal, but not ventral, hippocampal inputs. DLS(Pdyn) neurons inhibit LHA GABAergic neurons and confer context- and internal state-dependent calibration of feeding. Viral deletion of Pdyn in the DLS mimicked effects seen with optogenetic silencing of DLS Pdyn INs, suggesting a potential role for DYNORPHIN-KAPPA OPIOID RECEPTOR signaling in contextual regulation of food-seeking. Together, our findings illustrate how the dorsal hippocampus has evolved to recruit an ancient LHA feeding circuit module through Pdyn DLS inhibitory neurons to link contextual information with regulation of food consumption.
Keywords: GABA; context; dorsal hippocampus; dorsolateral septum; eating disorder; feeding; kappa opioid receptors; lateral hypothalamus; prodynorphin; somatostatin.
Conflict of interest statement
COMPETING INTERESTS None. DECLARATION OF INTERESTS The authors have no conflicts of interest to report
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources