

α-Synuclein oligomers form by secondary nucleation
- PMID: 39153989
- PMCID: PMC11330488
- DOI: 10.1038/s41467-024-50692-4
α-Synuclein oligomers form by secondary nucleation
Abstract
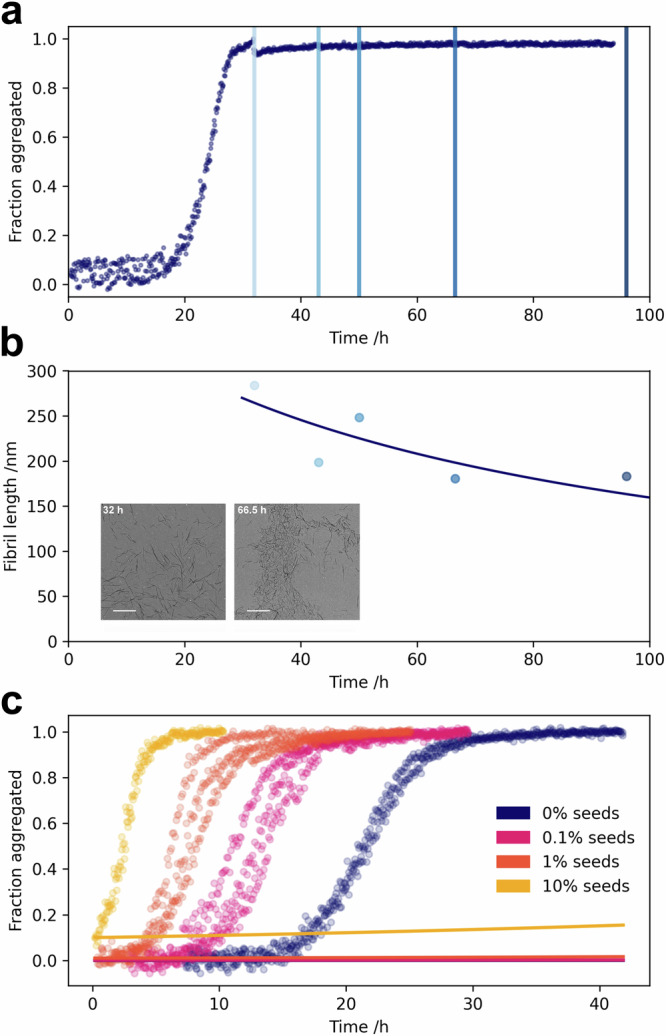
Oligomeric species arising during the aggregation of α-synuclein are implicated as a major source of toxicity in Parkinson's disease, and thus a major potential drug target. However, both their mechanism of formation and role in aggregation are largely unresolved. Here we show that, at physiological pH and in the absence of lipid membranes, α-synuclein aggregates form by secondary nucleation, rather than simple primary nucleation, and that this process is enhanced by agitation. Moreover, using a combination of single molecule and bulk level techniques, we identify secondary nucleation on the surfaces of existing fibrils, rather than formation directly from monomers, as the dominant source of oligomers. Our results highlight secondary nucleation as not only the key source of oligomers, but also the main mechanism of aggregate formation, and show that these processes take place under conditions which recapitulate the neutral pH and ionic strength of the cytosol.
© 2024. The Author(s).
Conflict of interest statement
At the time of initial submission, Georg Meisl and Alexander J Dear were employees of Wavebreak Therapeutics (formerly Wren Therapeutics). Michele Vendruscolo, Sara Linse, and Tuomas PJ Knowles are co-founders of Wavebreak Therapeutics (formerly Wren Therapeutics). The remaining Authors declare no competing interests.
Figures





References
MeSH terms
Substances
Grants and funding
- 101001615/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 European Research Council (H2020 Excellent Science - European Research Council)
- 841466/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 Marie Skłodowska-Curie Actions (H2020 Excellent Science - Marie Skłodowska-Curie Actions)
- 2015-00143/Vetenskapsrådet (Swedish Research Council)
LinkOut - more resources
Full Text Sources
